紅葉に象徴される葉の老化は、寿命の終着点ではありません。植物は季節や環境の変化を読み取り、自ら老化のタイミングを決定し、資源利用や生存戦略を最適化しています。1枚ずつ標識した2万枚以上の葉の追跡と分子解析から、老化が環境適応の鍵であることが見えてきました。
Ⅰ.はじめに:紅葉から見る葉の老化
暑い夏も終わり、山肌が少しずつ赤や黄に色づき始めました。秋の訪れを告げるこの紅葉は、誰もが一度は目にしたことのある風景でしょう。しかしこの美しい現象は、植物学の世界では「葉の老化」と呼ばれる生命現象の一つです。(図1)

では、なぜ植物は自分の葉を老化させ、枯らしてしまうのでしょうか。寿命が尽きたから? 役目を終えたから?それは半分正しく、半分間違いです。葉の老化の研究の歴史は古く100年ほど前から研究されてきました。その中で、葉の老化はただ受動的に起こるものではなく、植物が自らの判断で“積極的に”進めている現象であるということが明らかにされてきました。植物は環境の変化や季節の移ろいを敏感に読み取り、「この葉はもう役割を終えた」と判断した瞬間に、老化プログラムを起動します。 つまり、老化とは単なる“終わり”ではなく、植物が自らの成長と生存を最適化するための戦略なのです。この視点に立つと、これまで見慣れていた紅葉の景色が、まったく違う意味を持って見えてきます。
暑い夏も終わり、山肌が少しずつ赤や黄に色づき始めました。秋の訪れを告げるこの紅葉は、誰もが一度は目にしたことのある風景でしょう。しかしこの美しい現象は、植物学の世界では「葉の老化」と呼ばれる生命現象の一つです。(図1)
では、なぜ植物は自分の葉を老化させ、枯らしてしまうのでしょうか。寿命が尽きたから? 役目を終えたから?それは半分正しく、半分間違いです。葉の老化の研究の歴史は古く100年ほど前から研究されてきました。その中で、葉の老化はただ受動的に起こるものではなく、植物が自らの判断で“積極的に”進めている現象であるということが明らかにされてきました。植物は環境の変化や季節の移ろいを敏感に読み取り、「この葉はもう役割を終えた」と判断した瞬間に、老化プログラムを起動します。 つまり、老化とは単なる“終わり”ではなく、植物が自らの成長と生存を最適化するための戦略なのです。この視点に立つと、これまで見慣れていた紅葉の景色が、まったく違う意味を持って見えてきます。
Ⅱ.背景:落葉と常緑に共通する葉の老化の最適化原理
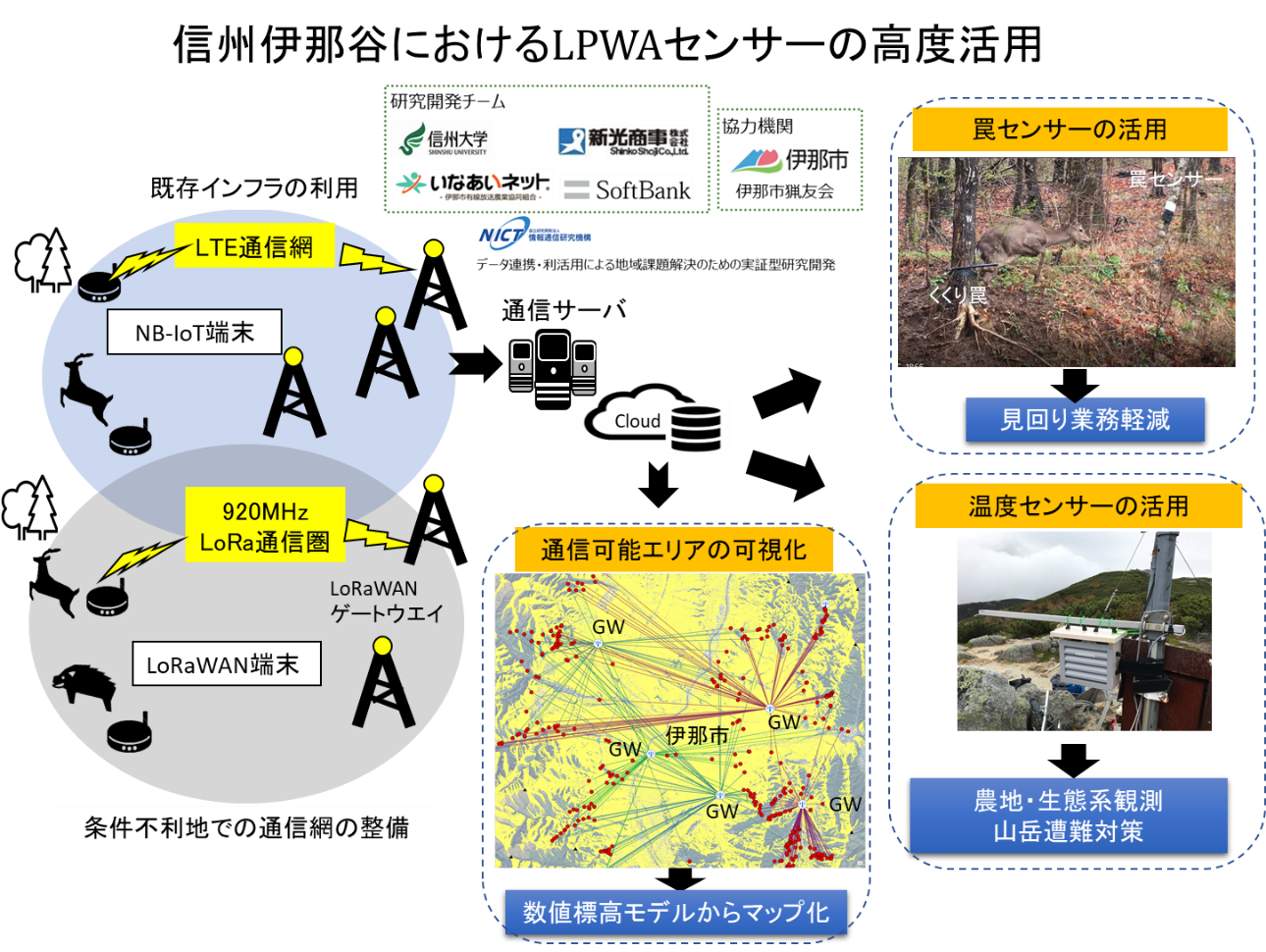
葉の老化は、植物の種類や生活史によってその姿を大きく変えます。たとえば落葉樹では、春に新しい葉が展開し、秋になると一斉に落葉するという明快なサイクルが見られます。一方で常緑樹は、一年中葉をつけているように見えますが、実際には新しい葉の展開と古い葉の老化が常に繰り返されているのです。 この違いは、植物が直面する”葉の生産コスト”と”光合成生産力”のバランスによって説明できます。葉は、まず一定のコストを払って作られ、その後は光合成によって生産活動を行います。しかし時間の経過とともに、生産力は少しずつ低下していきます。光合成産物の蓄積や活性酸素の蓄積などによって、光合成効率は次第に落ちていくのです。そのため、時間あたりの光合成効率は展葉直後は増加していき、やがて、減少に転じる瞬間が訪れます(図2左)。そのとき植物は、生産力が下がった葉から資源を回収し、捨てるという決断=老化を行い、個体全体として炭素獲得量を最大化します(図2右)。葉をただ枯らすのではなく、再利用できる栄養をしっかりと回収したうえで、新しい葉を展開するのです。
葉の老化は、植物の種類や生活史によってその姿を大きく変えます。たとえば落葉樹では、春に新しい葉が展開し、秋になると一斉に落葉するという明快なサイクルが見られます。一方で常緑樹は、一年中葉をつけているように見えますが、実際には新しい葉の展開と古い葉の老化が常に繰り返されているのです。 この違いは、植物が直面する”葉の生産コスト”と”光合成生産力”のバランスによって説明できます。葉は、まず一定のコストを払って作られ、その後は光合成によって生産活動を行います。しかし時間の経過とともに、生産力は少しずつ低下していきます。光合成産物の蓄積や活性酸素の蓄積などによって、光合成効率は次第に落ちていくのです。そのため、時間あたりの光合成効率は展葉直後は増加していき、やがて、減少に転じる瞬間が訪れます(図2左)。そのとき植物は、生産力が下がった葉から資源を回収し、捨てるという決断=老化を行い、個体全体として炭素獲得量を最大化します(図2右)。葉をただ枯らすのではなく、再利用できる栄養をしっかりと回収したうえで、新しい葉を展開するのです。
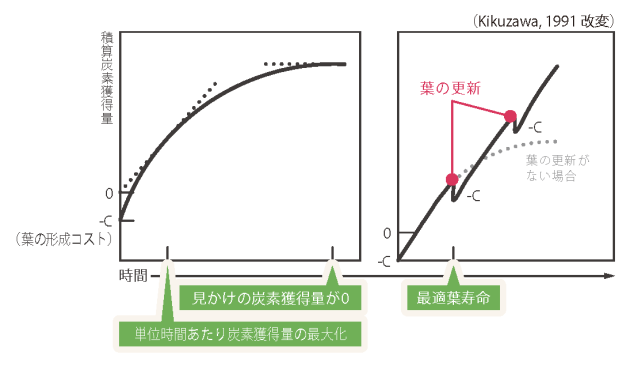
図2. 葉寿命の最適化
時間あたりの光合成による炭素獲得量は、葉の展開後に一度増加し、その後低下に転じる(左)
葉を適切な時点で更新することで、植物個体全体としての単位時間あたり炭素獲得量を最大化できる(右)
このような戦略的な老化の仕組みは、植物の生態にも深く関わっています。例えば、葉の生産コストが高く(=作るのが大変な葉)だったり、生産力が低く(=光合成の効率が悪い)環境では、植物はできるだけ長く葉を使い続ける必要があります。そのため、常緑樹は食害への防御コストが高い低緯度地域や、生産力が低くなる寒冷な高緯度地域で優勢になります。一方で、四季があるような中緯度地域では、春から秋に集中的に生産を行い、冬は休眠するという落葉樹のような「短期決戦型」の戦略が現れます。老化の仕組みは、植物がどこでどのように生きるかという“生態戦略”と深く結びついているのです。見た目の違いはあっても、落葉樹も常緑樹も、根底にある原理は同じです。
Ⅲ.多年生草本を用いた研究:季節に応じた老化制御の切り替えにいて
本研究では、常緑多年生植物 ハクサンハタザオ (Arabidopsis halleri subsp. gemmifera) を対象に、季節と老化制御の関係を詳細に解析しました。この植物は一年を通じて葉を展開・維持するため、生産期(春〜秋)から越冬期(秋〜冬)を経て翌春に至るまで、老化のプロセスを連続的に追跡できるという利点を持っています。さらに、モデル植物シロイヌナズナの近縁種であることから、豊富な遺伝情報を活用した分子解析が可能です。
私たちは、野外個体の葉を毎週標識してのべ20,000枚以上を追跡し、各葉の展開から老化・枯死に至るまでの運命を記録しました。その結果、老化のタイミングが季節によって根本的に異なることが明らかになりました。
生産期(春〜秋)には、葉は展開順に古いものから順次老化・枯死していきます。これは、古い葉の生産効率が低下すると植物が老化プログラムを起動し、古い葉を除去する戦略をとっているためです。一方、越冬期(秋〜冬)になると状況は一変します。老化は停止し、古い葉も若い葉も長期間維持され、翌年の繁殖期に葉齢にかかわらず一斉に枯死します。このことは、老化が単なる寿命ではなく、季節環境に応じて切り替えられる制御過程であることを示しています(図3)。
Ⅲ.多年生草本を用いた研究:季節に応じた老化制御の切り替えにいて
本研究では、常緑多年生植物 ハクサンハタザオ (Arabidopsis halleri subsp. gemmifera) を対象に、季節と老化制御の関係を詳細に解析しました。この植物は一年を通じて葉を展開・維持するため、生産期(春〜秋)から越冬期(秋〜冬)を経て翌春に至るまで、老化のプロセスを連続的に追跡できるという利点を持っています。さらに、モデル植物シロイヌナズナの近縁種であることから、豊富な遺伝情報を活用した分子解析が可能です。
私たちは、野外個体の葉を毎週標識してのべ20,000枚以上を追跡し、各葉の展開から老化・枯死に至るまでの運命を記録しました。その結果、老化のタイミングが季節によって根本的に異なることが明らかになりました。
生産期(春〜秋)には、葉は展開順に古いものから順次老化・枯死していきます。これは、古い葉の生産効率が低下すると植物が老化プログラムを起動し、古い葉を除去する戦略をとっているためです。一方、越冬期(秋〜冬)になると状況は一変します。老化は停止し、古い葉も若い葉も長期間維持され、翌年の繁殖期に葉齢にかかわらず一斉に枯死します。このことは、老化が単なる寿命ではなく、季節環境に応じて切り替えられる制御過程であることを示しています(図3)。

図3. 葉老化の季節変化
生産期(春〜夏)に展開した葉は短寿命であり、展開順に古い葉から順次枯死する(左上)
この過程では、下位に位置する古い葉が優先的に老化する(左下)
一方、越冬期(秋〜冬)に展開した葉は長寿命であり、冬季に老化はほとんど進行せず、春季の繁殖期に葉齢にかかわらず一斉に枯死する(右)
この制御は、外部刺激に対する応答にも現れます。生産期に葉を強制的に被陰させ、光環境を悪化させると老化が顕著に促進されるのに対し、越冬期では同じ刺激を与えても老化が誘導されず、処理間に差が見られないことがわかりました(図4)。 つまり、植物は季節を読み取り、「老化プログラムそのもののスイッチ」を環境に応じてオン・オフしているのです。 さらに、約150ほどの老化関連遺伝子を解析したところ、生産期と越冬後春期とでは老化の過程で発現が増加する遺伝子群が異なることが判明しました(図5)。 これは、植物が単に「老化を起こす・起こさない」を切り替えているだけでなく、季節ごとに異なる遺伝子プログラムを選択的に作動させていることを意味します。 これらの成果は、「常緑植物は年中ゆるやかに同じシステムで老化している」という従来の常識を覆すものです。常緑多年生植物であっても、老化は明確な季節性を持ち、異なる分子プログラムによって精密に制御されている──それが2万枚の葉を追跡した結果、初めて明らかになった真実です。
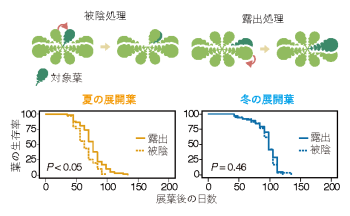
図4. 被陰処理による老化促進実験
糸による葉の位置操作により、対象葉を被陰または露出状態にした(上段)
生産期に展開した葉は被陰処理により生存率の低下が早まり、老化が促進された
一方、越冬期に展開した葉では処理間で有意な差は認められなかった(下段)
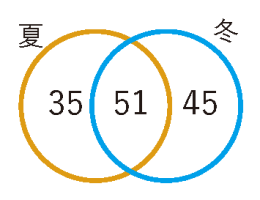
図5. 老化関連遺伝子の季節変化
老化過程で発現が増加する老化関連遺伝子のうち、
一部は季節を通じて共通していたが多くは生産期
および越冬後春季のいずれかで特異的に変化した
Ⅳ.将来展望:環境が違えば“最適な老化タイミング”も変化する
葉の老化は、生産と資源分配を最適化するための戦略であり、環境条件によって最適解は大きく変化します。今回明らかになった「生産葉から越冬葉への切り替えプログラム」は、現在の気候条件下で最適化された結果に過ぎません。標高、気温、日長などの環境が変化すれば、そのタイミングはズレる可能性があります。たとえば、高標高の厳しい環境では成長可能な期間が短くなるため、老化スケジュールはより早く、あるいはより長く引き延ばされる必要があるかもしれません。逆に、低標高では成長期間を最大限に活かす戦略へとシフトする可能性があります。 これまで行ってきたように単一の集団では「環境にどう応答するか」しか分かりませんが、異なる環境で進化した集団を比較することで、「どこまでが応答で、どこからが適応か」を明らかにすることが可能です。この視点は、気候変動の影響を考えるうえでも重要です。環境変化によって老化スケジュールがずれ、生産と資源再配分の最適化が崩れると、分布域や生存戦略そのものが変わる可能性があります。「葉がいつ老化するか」を理解することは、植物の分布・進化・未来を読み解く鍵でもあるのです。今後はこのような視点で研究を発展させていきたいと考えております。
Ⅴ.終わりに:研究するということ
私の研究は、「なんでもいいから自然をよく観察して、面白い現象を見つけろ」という指導から始まりました。最初は「低温で代謝が落ちるため、冬は葉の老化は止まるやろなぁ」と考えていました。しかしデータを重ねるうちに、その考えは覆されました。老化が止まるのは、寒さの“結果”ではなく、日長という環境の予兆を読み取り、冬が来る前にあらかじめ老化を停止していたのです。 自然は私たちの予想を超えます。データが出たあとは当たり前の結果なのですが、仮説とは基本的には当たりません。その驚きこそが、今も私を研究へと駆り立てています。山岳科学研究所の強みは、このような自然と向き合う研究を、野外調査から分子レベルの解析まで行えることです。 そして今、研究は「季節応答」の理解から「季節適応」の解明へと歩みを進めています。葉が枯れる──その当たり前の現象の裏には、驚くほど精密で戦略的な機構が隠されています。山の自然の中で植物を見つめ続けることで、私たちは生命が環境を“読む力”の本質に、少しずつ近づいているのです。
葉の老化は、生産と資源分配を最適化するための戦略であり、環境条件によって最適解は大きく変化します。今回明らかになった「生産葉から越冬葉への切り替えプログラム」は、現在の気候条件下で最適化された結果に過ぎません。標高、気温、日長などの環境が変化すれば、そのタイミングはズレる可能性があります。たとえば、高標高の厳しい環境では成長可能な期間が短くなるため、老化スケジュールはより早く、あるいはより長く引き延ばされる必要があるかもしれません。逆に、低標高では成長期間を最大限に活かす戦略へとシフトする可能性があります。 これまで行ってきたように単一の集団では「環境にどう応答するか」しか分かりませんが、異なる環境で進化した集団を比較することで、「どこまでが応答で、どこからが適応か」を明らかにすることが可能です。この視点は、気候変動の影響を考えるうえでも重要です。環境変化によって老化スケジュールがずれ、生産と資源再配分の最適化が崩れると、分布域や生存戦略そのものが変わる可能性があります。「葉がいつ老化するか」を理解することは、植物の分布・進化・未来を読み解く鍵でもあるのです。今後はこのような視点で研究を発展させていきたいと考えております。
Ⅴ.終わりに:研究するということ
私の研究は、「なんでもいいから自然をよく観察して、面白い現象を見つけろ」という指導から始まりました。最初は「低温で代謝が落ちるため、冬は葉の老化は止まるやろなぁ」と考えていました。しかしデータを重ねるうちに、その考えは覆されました。老化が止まるのは、寒さの“結果”ではなく、日長という環境の予兆を読み取り、冬が来る前にあらかじめ老化を停止していたのです。 自然は私たちの予想を超えます。データが出たあとは当たり前の結果なのですが、仮説とは基本的には当たりません。その驚きこそが、今も私を研究へと駆り立てています。山岳科学研究所の強みは、このような自然と向き合う研究を、野外調査から分子レベルの解析まで行えることです。 そして今、研究は「季節応答」の理解から「季節適応」の解明へと歩みを進めています。葉が枯れる──その当たり前の現象の裏には、驚くほど精密で戦略的な機構が隠されています。山の自然の中で植物を見つめ続けることで、私たちは生命が環境を“読む力”の本質に、少しずつ近づいているのです。