

生物多様性における世界的ホットスポットのなかのホットスポットである中部山岳地域
日本列島は「生物多様性のホットスポット」と表現されます。約2500万年前には、アジア大陸の東縁に位置していたとされる日本列島は、長い時間をかけて大陸から離裂することで列島を形成したとされます。いわゆる「大陸島」です。一方、小笠原諸島や伊豆諸島のように、海底火山が隆起して形成された「海洋島」的な要素も含まれています。海洋島の成立時には生物相は皆無であり、一方の大陸島でも離裂する際に入り込んだごくわずかな生物相からの遷移が始まることから、大陸島の生物相は「大陸の出店(サテライト)」であると喩えられます。しかしながら、日本列島の生物多様性は極めて高く、「世界的ホットスポット」とも言われますが、どうしてでしょうか?
様々な要因が考えられますが、以下に列記するような列島の立地や複雑な地史の影響を強く受けていると考えられます。
(1)アジアモンスーン気候の影響を受けて水が豊かで、森林がよく発達していること
(2)南北に長く複数のバイオーム(Biome, 注1)を縦断すること
(3)カムチャツカ(北東方),サハリン(北方)、朝鮮半島や中国本土(西方)、琉球列島を介して台湾や東南アジア地域(南方)など四方八方におよぶ複数の分散ルートの存在
(4)東日本と西日本がアジア大陸から独立した離裂(観音開き説: 図1)
(5)大きな4プレート(太平洋、フィリピン海、アジア、北米プレート)がぶつかりあうことによる激しい地殻変動
この列島上で生活するあらゆる生物たちは、多かれ少なかれ、これらの複雑な地史の影響を受けながら暮らしてきたはずです。そして、彼らの遺伝子には過去の歴史が刻み込まれているに違いない。このような背景から、「日本列島の複雑な地史と生物の系統進化を付き合わせて議論してみたい」想いを抱きました。
今なお、激しい地殻変動が生じている日本列島では、比較的若い地質年代での山岳形成などが生じてきました。古代の進化史を紐解くことよりも、比較的新しい時代の進化史の方が、より高い精度で紐解くことができます。つまり、日本列島は、地史と系統進化史の関連性を議論するには最適のフィールドであり、日本列島を舞台にした「生物系統地理」の研究は、日本列島ならではの「地の利」を活かした研究でもあるのです。
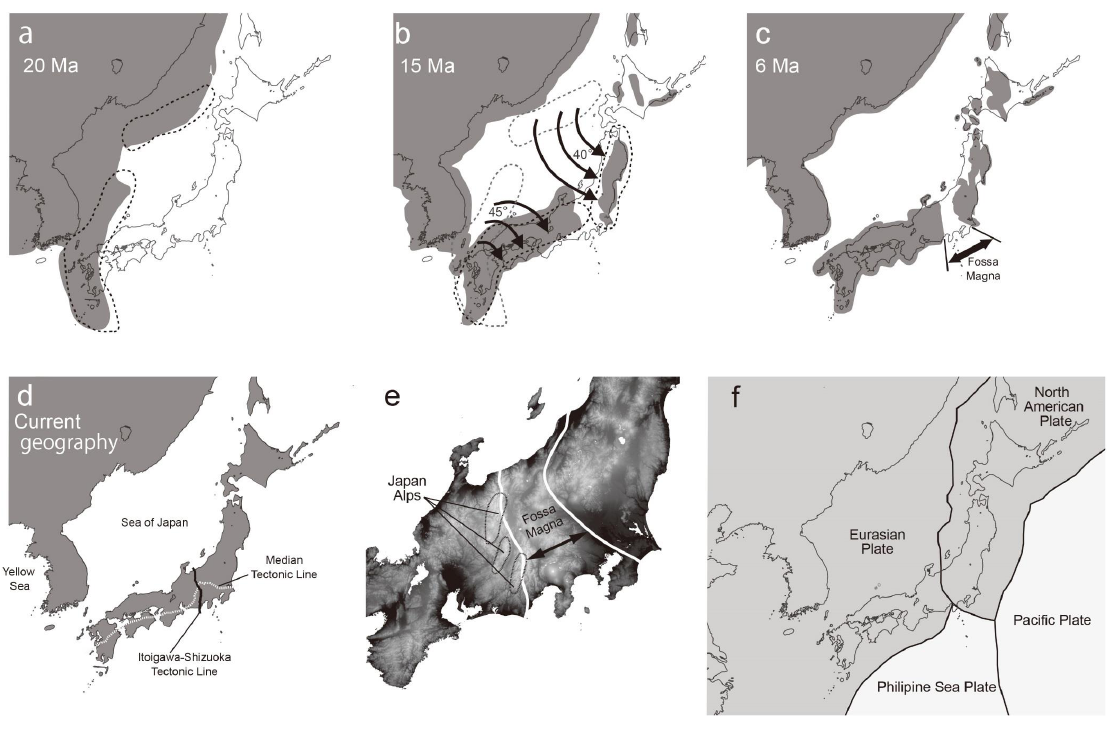
図1 日本列島の起源(観音開き説とフォッサマグナ)
日本列島はアジア大陸の東縁から、東日本と西日本が独立して観音開きのように離裂して形成されたとされる。東日本は半時計周り、西日本は時計周りの回転を伴ったことは、古地磁気の研究から明らかとなっている。また、5百万年前頃までの期間は、これらの境界域は海水面下であったとされる。このエリアは「フォッサマグナ(深い溝)」と呼ばれ、多くの生物種群において、東西日本での遺伝分化が認められることとも深く関連する。図はSaito and Tojo (submitted)より。
(注1) バイオーム(Biome)とは、気候的特性によって区分された地域(ツンドラ・熱帯降雨林など)に生息する生物群集の単位(生物群系)
山岳形成による河川争奪と水生昆虫の遺伝構造
〜かつては飛騨川へとながれていた上高地・梓川の水生昆虫のDNAからその痕跡を発見〜
上高地は信州の豊かな自然の象徴的存在です。3000m級の急峻な山岳から水を集めた梓川が上高地を貫流し、松本盆地へと流下しています。嶮しい連峰を背景に緩やかに流れる梓川のコントラストが上高地の魅力であるとも言われます。近年、山岳科学研究所の原山教授らの地質学的研究から、約1.2万年前まで、梓川は飛騨・高山川へとながれていたことが明らかとなりました。焼岳火山群の活動に因る造山活動あるいは山体崩壊により、(1)宮川-神通川(約64万年前以前)へとながれていた古梓川が、(2)高原川-神通川(約64〜1.2万年前)への流路変更を経て、現在のような松本方面へと流下するような(3)梓川-犀川-千曲(信濃)川への河川争奪が生じたとされます。(2-3)への過程では、古梓川が塞き止められ、巨大な自然のダム湖が形成されたとも考えられ、この湖底に堆積した石礫が現在の上高地の平らな地形の由来にもなっています。
このような地史は、当然のことながら、この地で暮らす生物種群にも大きく影響したはずです。そこで、これらのどの水系にも永年にわたって(少なくとも数十万年前には定着し)生息し、移動分散力が極めて弱いと考えられる水生生物・オビカゲロウ(カゲロウ目)を対象に、遺伝構造の解析を試みました。この結果、梓川の河川争奪に関係する3河川間で大きく遺伝的に分化していることが明らかとなりました。さらに、これらのいずれとも接続していたとされる上高地地域に生息しているオビカゲロウは、どの地域の遺伝構造をも複合的に持ち合わせていることが明らかとなりました(図2)。
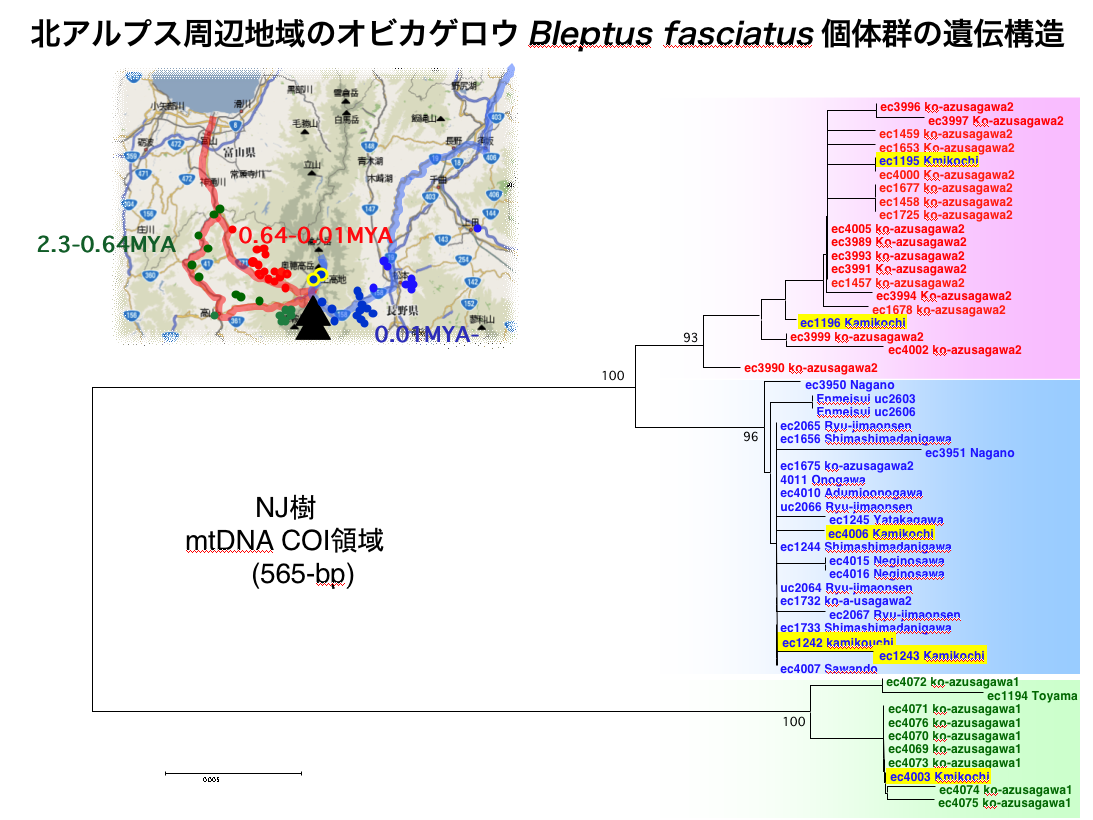
図2 オビカゲロウの系統地理
北アルプス南部(槍-穂高連峰)における河川争奪と遺伝分化の実例。河川源流域の細流に適応したオビカゲロウは、北アルプスの飛騨側(宮川、高原川)と松本側(梓川)とで大きく遺伝的に分化している(分子系統樹における配色と採取地点の配色は対応している)。上高地からは、いずれの系統も検出される。図はTojo et. al.(in prep.)より。
山岳形成が生物種内の遺伝的多様性を創出
〜山岳形成による集団間の遺伝分化〜
日本列島を含む東アジア地域を中心に、様々な動物種群における遺伝構造を解析してきた結果、地史と遺伝構造は見事なまでに合致していることが次々とあきらかになってきました。とくに河川に生息する昆虫類や淡水魚類を対象とした解析結果は目を見張るものがあります。河川で暮らす生き物たちは、線的ネットワークからなる水系内での移動分散を強いられますので、「面的」分散が可能な多くの生き物たちに対して、より分散性が低くなるためであると考えています。海峡形成や山岳形成による(種内)集団間での遺伝分化の事例も多数蓄積することができました(図3)。また、様々な生物種群での高精度の解析を蓄積することで、当初想定していたような「アジア大陸 → 日本列島」への分散(Downstream Dispersal)ばかりでなく、日本列島内で多様化した系統が再度アジア大陸へと分散するような「日本列島 → アジア大陸」への分散(Back Dispersal)も生じてきたことが明らかとなってきた。
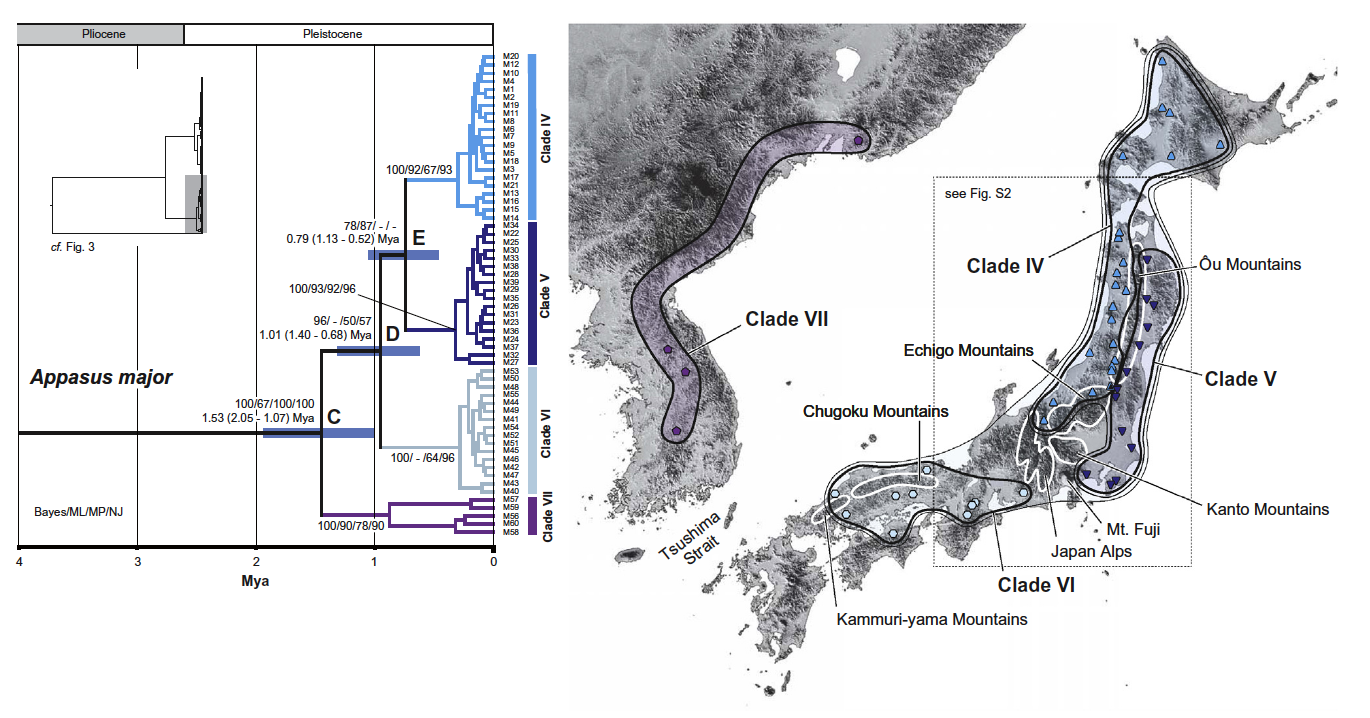
図3 オオコオイムシの遺伝構造と系統地理
対馬海峡による日本列島-アジア大陸間での遺伝分化、奥羽山脈の日本海側-太平洋側での遺伝分化、中部山岳域の東西での遺伝分化などが、顕著に認められる。図は Suzuki, Kitano and Tojo (2014) Mol. Phylogenet. Evol. より。
日本列島の山岳域は「進化生物学の実験室」
〜これからの展望〜
先にも述べたように、複雑な形成史をもつ日本列島、そして生物多様性のホットスポットである日本列島を舞台にした系統進化学・系統地理学は、多くのポテンシャルを有している。なかでも東日本と西日本の要素がぶつかりあう中部山岳域は、最重要地域と言える。山岳形成により稜線両側の山麓間での集団分化は、机上の理論としては古くから考慮されてきたものの、これらを詳説する事例などはほとんどなかったのが実状である。世界的にも稀な、今なおつづくドラスティックな地殻変動を有する日本列島の地の利を活かした研究成果を蓄積しつつ、世界的にもユニークな研究成果を信州からどんどん発信していきたいと考えています(例えば図4)。
これらの研究成果は、米国の分子系統進化学専門誌(Mol. Phylogetet. Evol.)や英国の生物学専門誌(Biol. J. Linn. Soc.)などに掲載されました(Suzuki et al., 2013, 2014; Sekine et al., 2010a, b, 2013; Kato et al., 2013など)。また本研究は、科学研究補助金・若手A(20687005)や挑戦的萌芽研究(23657064, 26650160)などのサポートを受けています。
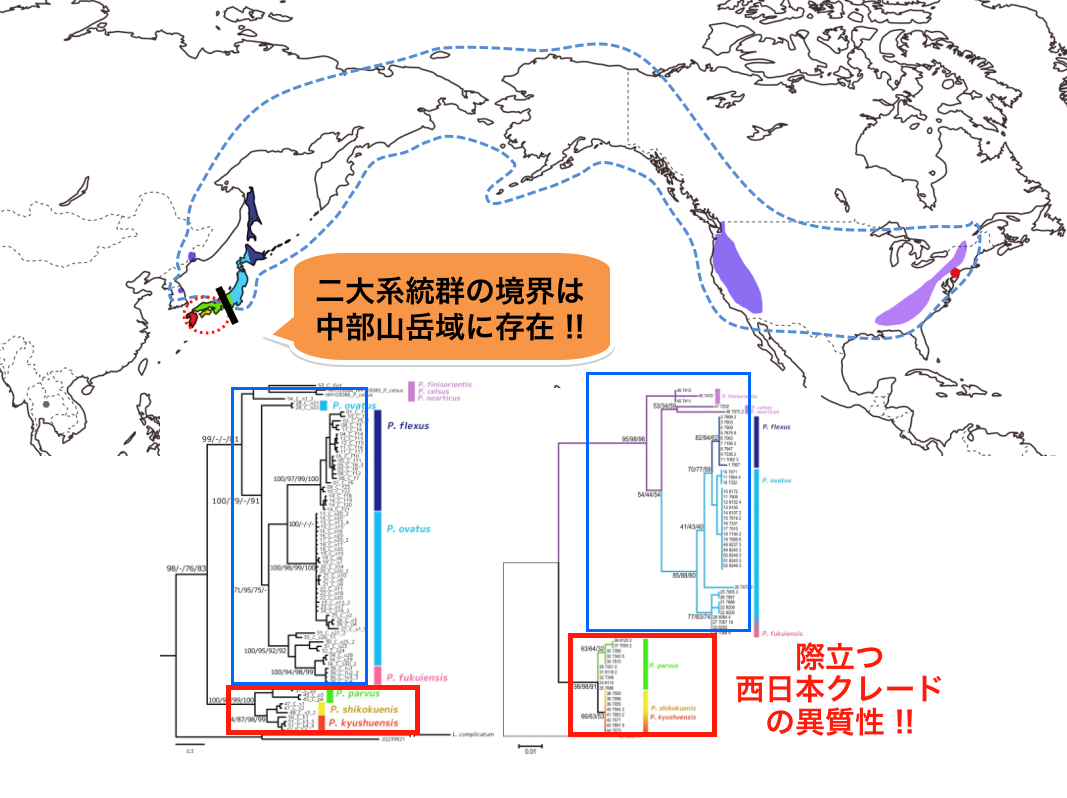
図4 カメノコヒメトビケラ類の分子系統地理
世界に9種生息するカメノコヒメトビケラ類全9種を用いた分子系統地理の解析結果。青色系統(1)北米大陸(アパラチア山脈・ロッキー山脈)+アジア大陸(ロシア沿海州)+サハリン+東日本の単系統性、赤色系統(2)西日本の単系統性が強く支持される。そしてこの世界スケールでの二大系統の境界が中部山岳地域に当たる。Araiya, Ito and Tojo (in prep.)より。

