研究トピックス
小笠原 寛 助教
(基盤研究支援センター 遺伝子実験支援部門)
 複合ストレスの乗り切り方
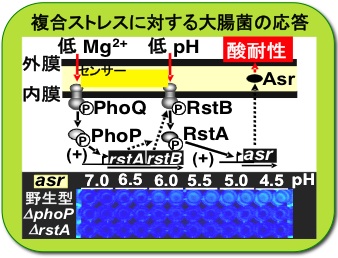
複合ストレスの乗り切り方
Mg2+の欠乏は細胞の生育の妨げとなり、そのような危機的条件下で加わるさらなる環境ストレスは細胞にとって脅威となります。Mg2+欠乏の危機に瀕した大腸菌が酸性条件に曝された際、それら複合的な条件を二種類のセンサーを使ってキャッチし、酸耐性遺伝子asrを働かせる仕組みを明らかにしました。
 大腸菌はどうやってバイオフィルムを形成するか?
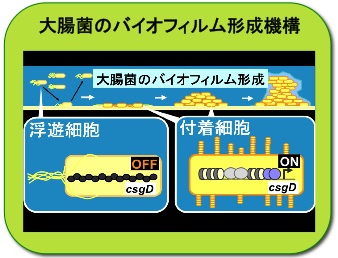
大腸菌はどうやってバイオフィルムを形成するか?
私たちの身近に存在する細菌の多くは、自然環境中ではバイオフィルムと呼ばれる共同体を作り棲息しています。腸内細菌科に属する大腸菌が、バイオフィルム形成に関わる遺伝子を働かせる仕組みについて調べ、csgD遺伝子のON/OFF切替が関係することを明らかにしました。
志田 敏夫 教授
 深海からポリマー産生菌の検索
深海からポリマー産生菌の検索
深海などの極限環境には未知の微生物が多数存在し、その中には人類にとって有用な物質を生産しているものもいます。それら微生物が作る新規のバイオファイバー・バイオマテリアルを解析し、利用する研究をしています。
 酵素反応の仕組みを解明
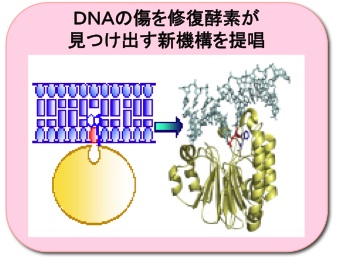
酵素反応の仕組みを解明
遺伝情報を担っている染色体DNAの傷をDNA修復酵素がどのようにして見つけ出して修復するか、遺伝子工学や蛋白質工学の手法を駆使して明らかにしました。酵素・基質複合体形成メカニズムに関して新知見が得られ、タンパク質の立体構造情報に基づく薬剤設計などへ応用できます。
下坂 誠 教授
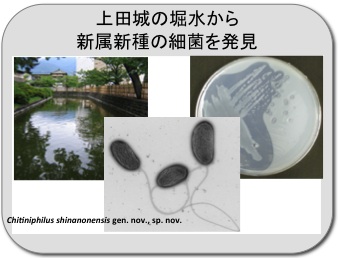 上田城の堀水から新属新種の細菌を発見
上田城の堀水から新属新種の細菌を発見
上田城の堀水から強力なキチン分解能力をもつ細菌を分離しました。種々の試験の結果、新属新種の細菌と判明し、学名 Chitiniphilus shinanonensis を付けて登録しました。現在、この細菌の強力なキチン分解酵素系について調査中です。
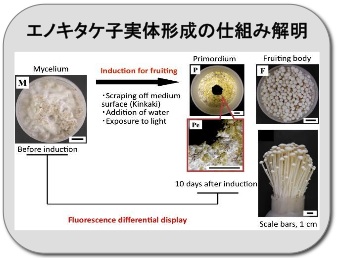 エノキタケ子実体形成の仕組みを解明
エノキタケ子実体形成の仕組みを解明
エノキタケ Flammulina velutipes のビン栽培のようすです。菌掻き、注水、光照射による子実体誘導処理によって子実体形成のスイッチが入ります。このとき、どのような遺伝子がはたらいているのか調べています。
高島 誠司 准教授
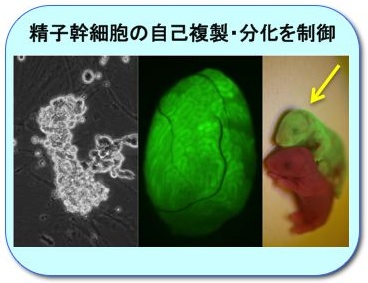 精子幹細胞の自己複製・分化を制御
精子幹細胞の自己複製・分化を制御
精子幹細胞 (GS細胞) の自己複製・分化のメカニズムを解明し、不妊治療や有用家畜の生産に応用します。(左) 試験管内で増殖するマウスGS細胞。(中) 緑色蛍光タンパクを発現するGS細胞を移植した精巣。緑色蛍光を発する精子ができている。(右) GS細胞由来の精子でできた仔マウス (矢印)。
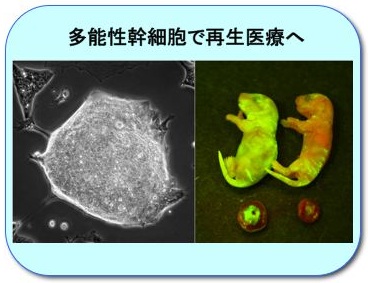 精子幹細胞の潜在的分化多能性を再生医療へ利用
精子幹細胞の潜在的分化多能性を再生医療へ利用
精子幹細胞はごく稀に脱分化し、iPS細胞と同様の分化多能性を持つ「多能性精子幹細胞 (mGS細胞)」に変化します。この脱分化機構を自在に制御することで、再生医療への応用を目指します。(左) GS細胞から脱分化して生じたmGS細胞。(右) mGS細胞からなるキメラマウス (緑色蛍光部はmGS細胞由来)。
田口 悟朗 教授
 植物の香り物質を探して!
植物の香り物質を探して!
植物は生理活性物質や香り物質など、さまざまな有用物質を作り出します。それらの物質が植物中でどのように生合成されるのかを明らかにするとともに、そこで得られた知見をもとに、例えば、香りの良い花を作りだす取り組みを進めています。
 ヒカリゴケの培養に成功!
ヒカリゴケの培養に成功!
ヒカリゴケは山地の洞窟などに希に自生しており、緑色の反射光を出すことで知られていますが、自然開発によって絶滅が危惧されています。このヒカリゴケを試験管内培養することに成功しました。培養後に土の上で生育させると写真のような光を放ちます。
根岸 淳 助教
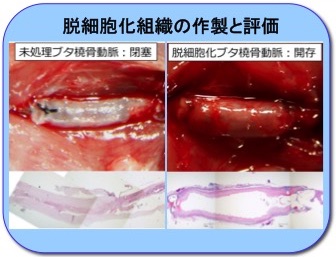 脱細胞化組織の作製と機能解析
脱細胞化組織の作製と機能解析
生体組織から細胞を除去した脱細胞化組織を作製し、その生体反応を解析し、医療用材料および生体機能解明のツールとして応用します。写真左はラット頸動脈移植3日目の未処理ブタ橈骨動脈、写真右はラット頸動脈移植2週間目の脱細胞化ブタ橈骨動脈です。脱細胞処理によって異種移植の免疫拒絶を回避し、血管として機能しています。
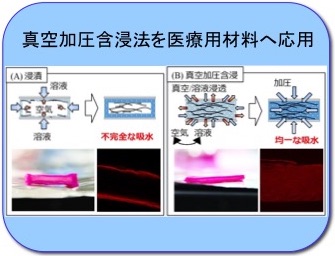 真空加圧含浸法の医療用材料への応用
真空加圧含浸法の医療用材料への応用
金属加工、食品加工分野で使用されている真空加圧含浸法の医療用材料への応用展開を検討しています。凍結乾燥した脱細胞化組織を溶液に浸漬すると組織内の空気が抜けにくいため不均一な溶液浸透になります (左図A) が、真空加圧含浸法により組織内の空気を除去した状態で溶液導入が可能になります (左図B)。
野川 優洋 准教授
 非病原性に変異させたアグロバクテリウムを作製
非病原性に変異させたアグロバクテリウムを作製
アグロバクテリウム菌は植物細胞に菌のDNAを運びクラウンゴール病を引き起こします。そこでトランスポゾン挿入変異法により病原性がなくなった変異菌を作り、これらの変異菌からアグロバクテリウムの病原性に関わる新規遺伝子として「abvA遺伝子」を見いだしました。
 植物の効率的な形質転換
植物の効率的な形質転換
クラウンゴール病 (写真左) を引き起こす病原菌として知られている Agrobacterium tumefaciens が植物細胞の核にDNAを運ぶ性質を利用し、短期間に優良作物を作出することができます。これにより、収量が増大したソバ、茎が太くなったケナフ、枝分かれの増えたクワ (写真右)、などの作出に成功しました。
野村 隆臣 准教授
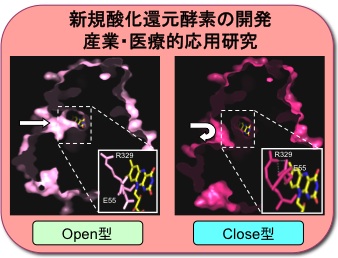 新規酸化還元酵素の開発
新規酸化還元酵素の開発
新規酸化還元酵素であるグリシンオキシダーゼについて、基質阻害をもたらす要因を調べることで酵素特性や触媒メカニズムを解明しようとしています。遺伝子工学的技術によってさらなる機能改良を加えることで産業・医療的に展開できる応用研究を進めます。
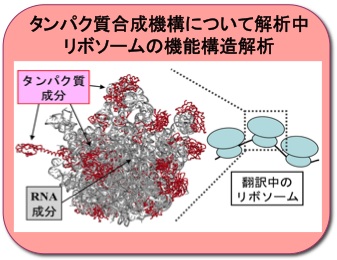 リボソーム中のタンパク質の役割解明
リボソーム中のタンパク質の役割解明
生体内タンパク質合成(翻訳)装置である「リボソーム」は数種のRNAと数十種のタンパク質成分より構成される超分子集合体です。リボソームの特徴的な突起構造を形成するタンパク質成分が翻訳伸長因子の選択性に直接関与することを「異種生物間ハイブリッドリボソーム」を構築することにより明らかにしました。
保地 眞一 教授
 ラットの顕微授精技術を確立
ラットの顕微授精技術を確立
鎌状の形態をしたラット精子頭部を顕微鏡下で卵子内に注入 (ICSIという) し、代理母に移植して産仔を得る技術を確立しています。さらにフリーズドライ処理で粉末にした精子を冷蔵庫で一年間保存し、ICSI後に産仔発生例を得ました。
 ウシ受精卵の凍結保存技術を確立
ウシ受精卵の凍結保存技術を確立
体外受精技術や顕微授精技術を駆使すれば、ウシの胚盤胞を実験室環境下で作製することができます。これらを超急速冷却ガラス化保存法という方法で凍結保存することにより、融解後でもほぼ100%蘇生させられるようになりました。
松村 英夫 准教授
(基盤研究支援センター 遺伝子実験支援部門)
 植物の性はどうやって決まるのか?
植物の性はどうやって決まるのか?
植物には雌雄異株のもの、同じ個体で雌雄異花のもの、両性花のものなど、非常に多様な性の様式が知られています。とくに果実生産に影響を与える農作物 (野菜、果樹) について、それらの性決定メカニズムを調べるための研究を進めています。
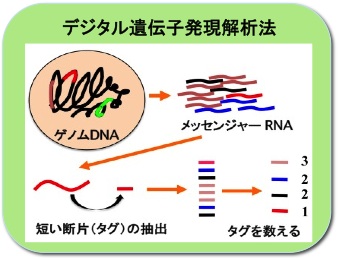 遺伝子の発現をデジタルデータで見る
遺伝子の発現をデジタルデータで見る
生物は数千から数万種類の遺伝子 (DNA) を持っています。環境や生長に応じて必要な遺伝子からmRNAが作られ、それを基にタンパク質が作られます。これらmRNAを一度に数十万本以上数えられる方法を開発し、どの遺伝子がどんな時にどれくらいコピーされているかをデジタルデータで知ることができました。
![]()