高山帯の「短い夏」を生き抜く秘訣 カギは休眠の有無!?
― 山岳域での生物多様性創生に関わる「遺伝的分化」と「生態的分化」の関係性を究明 ―
2024年6月13日
【研究成果のポイント】
・乗鞍岳の高山帯・亜高山帯で,サハリントビケラの生活史調査を実施しました
・標高差約1,000 mの高山帯・亜高山帯では生息環境に大きな差異がありました
・高山帯・亜高山帯のサハリントビケラの生活史にも大きな差異が検出されました
・高山帯では幼虫の発育開始が遅れ,成虫期の「夏眠」が欠如していました
・標高帯間で卵成熟・交尾時期に差異がみられ,生殖隔離(繁殖期のズレ)が生じていました
・生活史のズレによる生殖隔離は,高山帯・亜高山帯での遺伝的分化の要因の可能性があります
・高山帯への適応と発現遺伝子の関係性について,ゲノムワイドな解析を展開中です(今後の課題)
・乗鞍岳の高山帯・亜高山帯で,サハリントビケラの生活史調査を実施しました
・標高差約1,000 mの高山帯・亜高山帯では生息環境に大きな差異がありました
・高山帯・亜高山帯のサハリントビケラの生活史にも大きな差異が検出されました
・高山帯では幼虫の発育開始が遅れ,成虫期の「夏眠」が欠如していました
・標高帯間で卵成熟・交尾時期に差異がみられ,生殖隔離(繁殖期のズレ)が生じていました
・生活史のズレによる生殖隔離は,高山帯・亜高山帯での遺伝的分化の要因の可能性があります
・高山帯への適応と発現遺伝子の関係性について,ゲノムワイドな解析を展開中です(今後の課題)
【背景】
山岳では1,000 mの標高差で気温が約6 ℃変化するように,標高により環境はダイナミックに変化する.また,高山帯は極端な低温や乾燥に晒され,生物にとっては過酷な環境といえる.高山帯に棲息する生物種は,このような環境に適応しながら生きている.今回,乗鞍岳の高山帯・亜高山帯に棲息する寒冷適応した水生昆虫の一種,サハリントビケラAsynarchus sachalinensis Martynov, 1914を対象に,ほぼ全ての生活史をカバーする2022年4月から11月までの8か月間に渡り,毎週の現地での詳細な調査を実施した.その結果,高山帯・亜高山帯のいずれにおいても年一化(1年1世代)であるものの,標高帯間で幼虫の発育時期,成虫の出現時期には大きな差異(ズレ)が示された.特に,高山帯に棲息する集団は亜高山帯の集団に比べて幼虫の発育開始時期が大幅に遅れ,成虫では低地の集団のサハリントビケラで一般的に観察される夏の高温・乾燥を乗り切るための休眠(夏眠)が観察されなかった.このような生活史は夏が短い高山帯で生存し,世代交代を繰り返すことに適した生態的特徴であると推察される.さらに,高山帯と亜高山帯の集団間では,成虫の活動時期や交尾時期に違い(ズレ)がみられたことから,同じ山塊の高山帯と亜高山帯の集団間であっても交配は起こりにくく,遺伝的交流が生態的にも制限されている可能性が示唆された.実際に,高山帯・亜高山帯の遺伝系統は大きく分化しており(我々による先行研究; Suzuki et al., 2024)やゲノムワイドな遺伝子解析においても高山帯・亜高山帯での遺伝子流動は全く検出されない.ひとつづきの山塊である乗鞍岳の高山帯・亜高山帯における系統の分化(生殖的隔離が成立しているとなれば種分化にも繋がる分化)はとても興味深い現象であり,山岳域における生物多様性創出メカニズムを追究する上での重要な知見といえる.
山岳では1,000 mの標高差で気温が約6 ℃変化するように,標高により環境はダイナミックに変化する.また,高山帯は極端な低温や乾燥に晒され,生物にとっては過酷な環境といえる.高山帯に棲息する生物種は,このような環境に適応しながら生きている.今回,乗鞍岳の高山帯・亜高山帯に棲息する寒冷適応した水生昆虫の一種,サハリントビケラAsynarchus sachalinensis Martynov, 1914を対象に,ほぼ全ての生活史をカバーする2022年4月から11月までの8か月間に渡り,毎週の現地での詳細な調査を実施した.その結果,高山帯・亜高山帯のいずれにおいても年一化(1年1世代)であるものの,標高帯間で幼虫の発育時期,成虫の出現時期には大きな差異(ズレ)が示された.特に,高山帯に棲息する集団は亜高山帯の集団に比べて幼虫の発育開始時期が大幅に遅れ,成虫では低地の集団のサハリントビケラで一般的に観察される夏の高温・乾燥を乗り切るための休眠(夏眠)が観察されなかった.このような生活史は夏が短い高山帯で生存し,世代交代を繰り返すことに適した生態的特徴であると推察される.さらに,高山帯と亜高山帯の集団間では,成虫の活動時期や交尾時期に違い(ズレ)がみられたことから,同じ山塊の高山帯と亜高山帯の集団間であっても交配は起こりにくく,遺伝的交流が生態的にも制限されている可能性が示唆された.実際に,高山帯・亜高山帯の遺伝系統は大きく分化しており(我々による先行研究; Suzuki et al., 2024)やゲノムワイドな遺伝子解析においても高山帯・亜高山帯での遺伝子流動は全く検出されない.ひとつづきの山塊である乗鞍岳の高山帯・亜高山帯における系統の分化(生殖的隔離が成立しているとなれば種分化にも繋がる分化)はとても興味深い現象であり,山岳域における生物多様性創出メカニズムを追究する上での重要な知見といえる.

【研究対象種,対象地域と方法について】
サハリントビケラAsynarchus sachalinensis Martynov, 1914は,本州中部山岳域を分布の南西限に,東北地方や北海道の山岳域,ロシア・サハリンに分布・棲息するトビケラ※1の仲間である(図1).寒冷適応した水生昆虫の一種であり,中部山岳域の高山帯のように,寒冷で夏が極めて短い環境にも適応している.こうした高山帯に棲息するサハリントビケラは,亜高山帯とは異なる生態的特徴をもつことが予想されてきた.
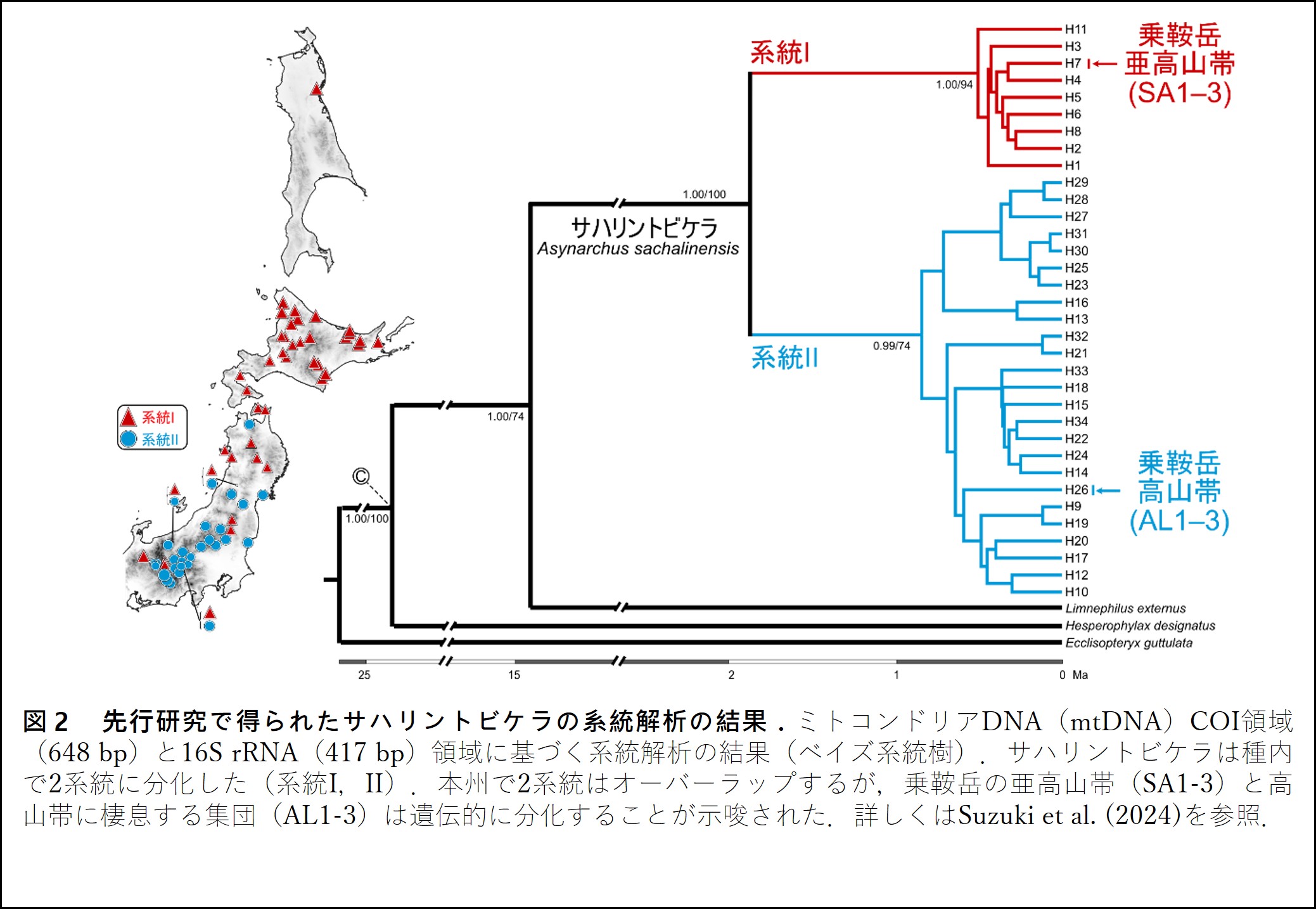
本種を対象とした我々の以前の研究では,mtDNA COI,16S rRNA領域に基づく系統解析により,遺伝的に大きく分化した2つの種内系統の存在を究明した(Suzuki et al., 2024).そして,これら種内系統は本州で分布域が広くオーバーラップするものの,標高帯により,棲息場所が分化している傾向がみられ,棲息環境と遺伝構造との関連性が示唆された.また,長野県・岐阜県にまたがる乗鞍岳(標高:3,026 m)にも本種は分布しており,標高2,700 m付近の高山帯と標高1,300 m付近の亜高山帯に棲息する集団は,遺伝的に大きく分化していることも示唆された(図2).
サハリントビケラAsynarchus sachalinensis Martynov, 1914は,本州中部山岳域を分布の南西限に,東北地方や北海道の山岳域,ロシア・サハリンに分布・棲息するトビケラ※1の仲間である(図1).寒冷適応した水生昆虫の一種であり,中部山岳域の高山帯のように,寒冷で夏が極めて短い環境にも適応している.こうした高山帯に棲息するサハリントビケラは,亜高山帯とは異なる生態的特徴をもつことが予想されてきた.
本種を対象とした我々の以前の研究では,mtDNA COI,16S rRNA領域に基づく系統解析により,遺伝的に大きく分化した2つの種内系統の存在を究明した(Suzuki et al., 2024).そして,これら種内系統は本州で分布域が広くオーバーラップするものの,標高帯により,棲息場所が分化している傾向がみられ,棲息環境と遺伝構造との関連性が示唆された.また,長野県・岐阜県にまたがる乗鞍岳(標高:3,026 m)にも本種は分布しており,標高2,700 m付近の高山帯と標高1,300 m付近の亜高山帯に棲息する集団は,遺伝的に大きく分化していることも示唆された(図2).
本研究は,「サハリントビケラの高山帯への生態的な適応がどのように生じてきたのか?」,「高山帯の環境に適応するような生態的特徴が,遺伝的分化を維持・促進する要因となりえるのか?」といった疑問のもと,2022年に乗鞍岳の高山帯・亜高山帯のそれぞれ3か所の池沼に棲息する集団を対象に,幼虫の発育ステージと成虫の出現パターンを,毎週の詳細な調査により究明した(高山帯:5月24日~10月18日 の計21回,亜高山帯:4月10日~11月17日 の計32回)(図3).

【結果・考察】
水温と幼虫の発育ステージの差異
幼虫の発育ステージと成虫の出現パターンから,高山帯・亜高山帯いずれの集団も化性※2は年一化であることが明示された.亜高山帯では,早春から幼虫が発育を開始し,初夏には蛹が観察されたのに対し,高山帯は初夏より幼虫の発育が開始し,盛夏に蛹化が観察された.つまり,幼虫の発育の開始時期は約1ヶ月も高山帯の方が遅れていた.これらの幼虫の発育プロセスは,データロガーを用いて計測された各調査地の水温変動のデータとも深く関係していた(図4).高山帯の3つの調査地間に,融雪時期(水温が上昇する時期)に違い(ズレ)がみられ,それに応じてた幼虫の発育時期にも差がみられた(図4).
水温と幼虫の発育ステージの差異
幼虫の発育ステージと成虫の出現パターンから,高山帯・亜高山帯いずれの集団も化性※2は年一化であることが明示された.亜高山帯では,早春から幼虫が発育を開始し,初夏には蛹が観察されたのに対し,高山帯は初夏より幼虫の発育が開始し,盛夏に蛹化が観察された.つまり,幼虫の発育の開始時期は約1ヶ月も高山帯の方が遅れていた.これらの幼虫の発育プロセスは,データロガーを用いて計測された各調査地の水温変動のデータとも深く関係していた(図4).高山帯の3つの調査地間に,融雪時期(水温が上昇する時期)に違い(ズレ)がみられ,それに応じてた幼虫の発育時期にも差がみられた(図4).
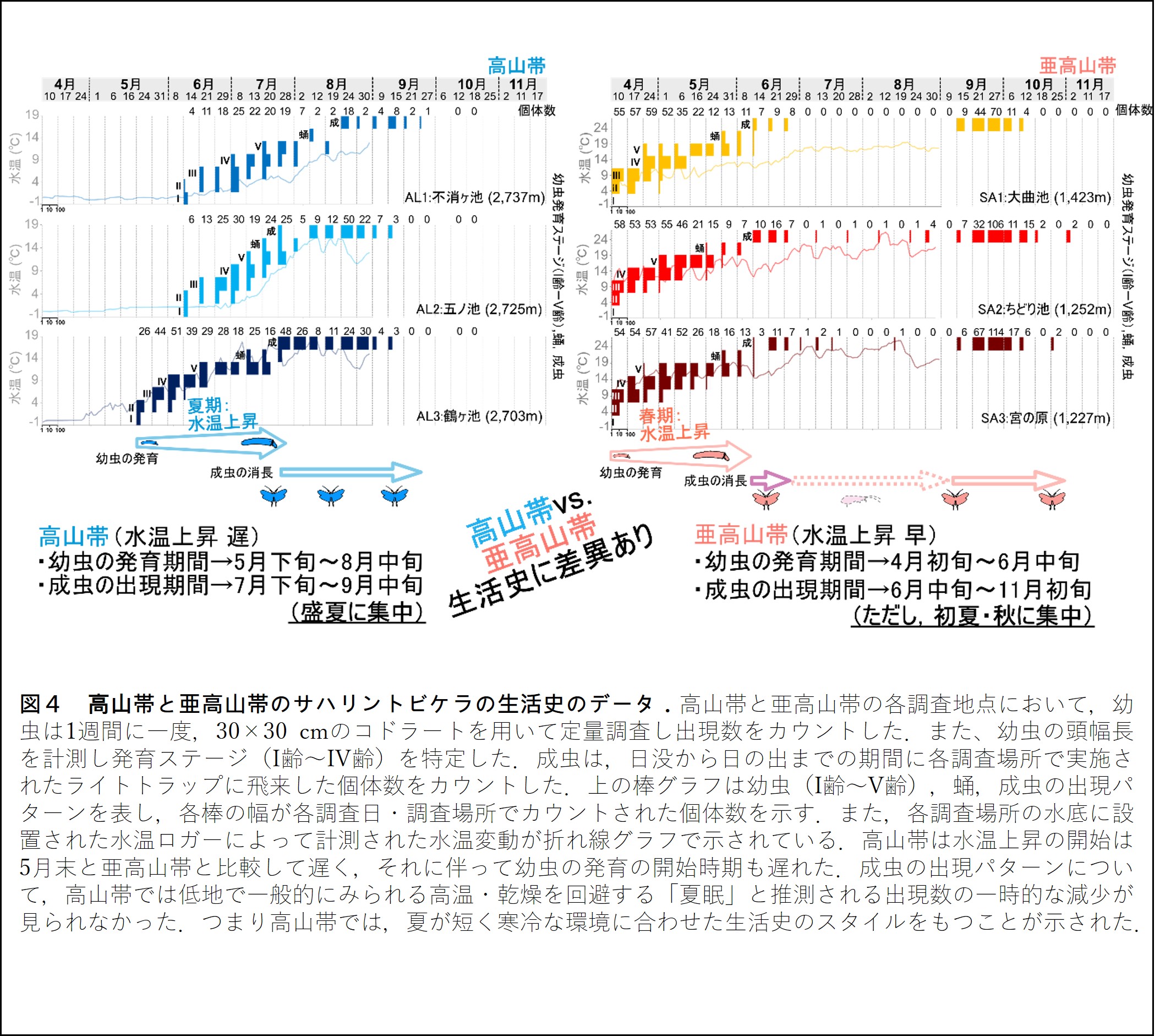
棲息環境の水温は,水生昆虫の成長・発育パターンを規定する要因の一つである.乗鞍岳は盛夏になっても残雪が多く,調査水域にも8月頃まで積雪がみられた.水域を覆う積雪により水温上昇が遅れ,それに伴い幼虫の成長・発育が遅れたと推察される.同一山塊の高山帯と亜高山帯の間では,直線距離では約10 kmしか離れていない.しかし,1,000 m以上ある標高差は気候的環境(温度・降雪量など)に大きな差異をもたらし,幼虫の成長・発育に影響を及ぼしたと考えられる.
高山帯と亜高山帯には,大きな環境の差異があり,特に高山帯に棲息する昆虫にとって,活動に適した限られた期間に繁殖し,確実に子孫を残す必要がある.では,「どのような方法で高山帯の厳しい環境に適応し,子孫を残しているのか?」,成虫の生活史※3パターンを調べることでその糸口を追究した.
生息環境と夏眠(卵巣休眠)の有無
成虫(オス,メス,卵巣未成熟メス)の出現パターンを高山帯・亜高山帯で比較したところ,高山帯では7月末から9月中旬にかけて成虫が集中的に観察されたのに対し,亜高山帯では6月中下旬にかけて観察された後,7,8月はほとんど観察されず,9月中旬から10月中旬にかけて再び観察された(図4).
高山帯と亜高山帯には,大きな環境の差異があり,特に高山帯に棲息する昆虫にとって,活動に適した限られた期間に繁殖し,確実に子孫を残す必要がある.では,「どのような方法で高山帯の厳しい環境に適応し,子孫を残しているのか?」,成虫の生活史※3パターンを調べることでその糸口を追究した.
生息環境と夏眠(卵巣休眠)の有無
成虫(オス,メス,卵巣未成熟メス)の出現パターンを高山帯・亜高山帯で比較したところ,高山帯では7月末から9月中旬にかけて成虫が集中的に観察されたのに対し,亜高山帯では6月中下旬にかけて観察された後,7,8月はほとんど観察されず,9月中旬から10月中旬にかけて再び観察された(図4).
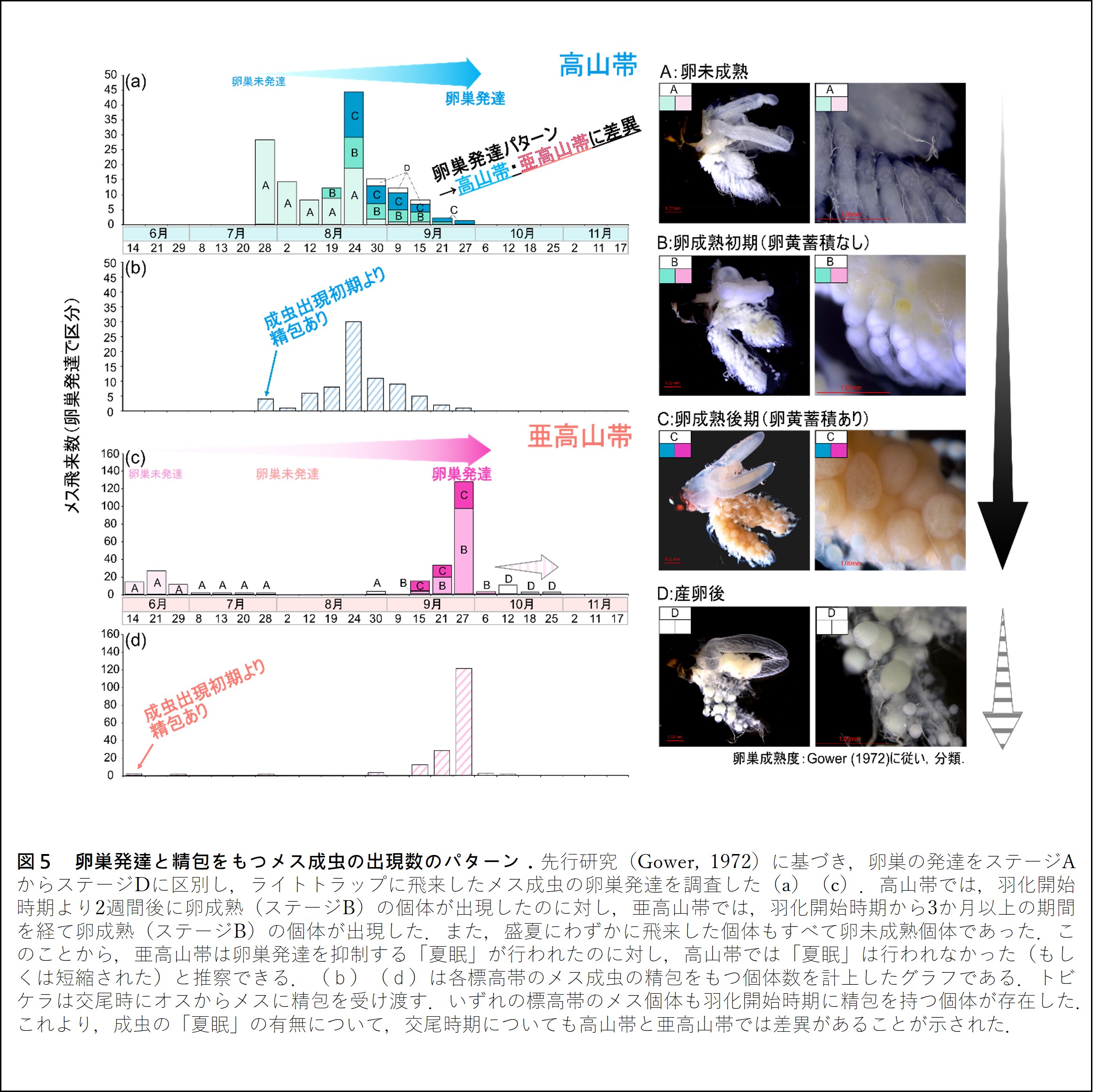
また,観察されたメスについて,卵成熟を先行研究に準じて4つのステージ(ステージA:卵未成熟期,ステージB:卵黄が観察されない時期,ステージC:卵黄が観察される成熟卵をもつ時期,ステージD:産卵後)に区分し,卵成熟の時期と程度を標高帯間で比較したところ,高山帯における卵成熟個体は,高山帯で成虫が初めて観察された日から約2週間後に観察された.一方,亜高山帯では,卵成熟個体は成虫が初めて観察されてから約3か月後の秋に観察され,盛夏にわずかに飛来した成虫は,いずれも卵未成熟のメスであった(図5).つまり,成虫の出現・卵成熟のパターンも高山帯・亜高山帯の間で大きな差異(大きなズレ)があった.
亜高山帯における生活史調査の結果,亜高山帯では成虫が盛夏に「夏眠(卵巣休眠)」している可能性が示唆された.「夏眠」とは,夏季において高温や乾燥に耐えるように代謝を一時的に低下(もしくは停止)させる休眠パターンであり,サハリントビケラを含む近縁なトビケラ類においても成虫期に夏眠し,卵巣発達を一時的に停止させる,いわゆる「卵巣休眠」を行うことが知られている.サハリントビケラの生活史について追究した先行研究では,初夏に出現した成虫は,盛夏に出現頻度は極めて減少し,再び秋に出現頻度が増加するとしている.北米に棲息する同属別種においても,同様の傾向が知られている.これらのことから,夏眠を有する生活史はサハリントビケラの一般的な生活史パターンであると推察される.夏眠を有する生活史パターンは,従来,寒冷な環境に棲息していたサハリントビケラが,高温や乾燥にさらされる可能性が高い地域に分布域を拡大し,環境に適応した結果,獲得した形質と考えられる.
一方,高山帯は亜高山帯とは異なり,成虫が集中的に出現し,卵未成熟の成虫が出現する期間は亜高山帯と比べて短期間であった.従って高山帯の集団には,夏眠期間が「ない」もしくは「短縮」されている可能性が示された.本研究によって示唆された,高山帯に棲息する集団の夏眠の欠如(もしくは短縮)する生活史パターンは,活動に適する期間が短い地域で繁殖を成功させる可能性の高い生態的特徴であると推察される.ノルウェーなど北極圏に近い地域に分布するサハリントビケラの近縁種の中には夏眠を行わない種があることも知られている.高緯度地域,高山帯はいずれも寒冷で夏が短い.こうしたことから,気温や日長などの要因が複合的に機能し,成虫の夏眠の有無に寄与していると考えられる.
以上より,サハリントビケラが棲息環境に応じて異なる適応をしていることが示唆された.次に,生活史の違いと標高帯間の遺伝的分化との関連性について追究した.
生息環境と夏眠(卵巣休眠)の有無
サハリントビケラは,オスが精子の入った袋「精包」をメスへと渡すことで交尾が完了する.つまり,メスの腹部にある精包の有無を調べることで,交尾時期を知ることができる.それぞれの標高帯で精包の有無を確認したところ,いずれの標高帯においても,成虫が観察され始めた直後(高山帯:7月下旬,亜高山帯:6月中旬)に,卵が未成熟であるメスの体内から精包が確認された(図4).このことは,本種は羽化直後の状態で,既に交尾を済ませており,また,それぞれの標高帯では交尾時期が異なる(ズレている)ことも示している.また,夏眠の有無の違いにより,成虫の活動時期も標高帯間で分化していることから,それぞれの集団間で交配が生じる可能性は低い.したがって,集団間の交配前生殖隔離が働いており,集団間の遺伝的分化が維持・促進されていると推測される.現在,標高帯間に存在する遺伝子流動の程度(遺伝的障壁の大きさ)を評価するべく,ゲノムワイドに存在する一塩基多型(SNPs)※4情報に基づく遺伝子解析を進めている.
結論
本研究は,同一山塊内の高山帯・亜高山帯という極めて近接する集団を形成する水生昆虫を対象に,生活史の差異(ズレ)を明示し,それぞれの生活史が異なる環境下において適応的な意義をもつ可能性を示した初めての事例である.今後,より詳細な遺伝子解析の結果を加味し,標高帯間に存在する遺伝的障壁を明らかにしていく予定である.
将来的な研究として,「生活史の差異は遺伝的に固定化されているのか?」,また「それぞれの環境への適応に関与する遺伝子は存在するのか?」といったような疑問を追究すべく,飼育実験やゲノムワイド関連解析(GEAS)※5に発展させ,山岳における生物多様性創出メカニズムの解明に前進していくことを目指す.
【用語解説】
※1 トビケラ: 毛翅目の昆虫類(鱗翅目の蝶や蛾に近縁な昆虫のグループ)で,幼虫は芋虫型,ほとんどの種が水中で生活する.口から吐く糸で筒状の巣(筒巣)を形成し,巣内で生活する種も多い.成虫は陸上生活するが,翅には(チョウ類やガ類がもつような)鱗粉ではなく,細かな毛が生えている.
※2 化性: 昆虫が1年間に世代を繰り返す性質のこと.1年間に1世代が繁殖し,一年周期で世代が繋がるような生活史をもつことを年1化(あるいは,単に「年一世代」)という.
※3 生活史: 生物が生まれてから死ぬまでの過程.昆虫の場合,卵,幼虫,蛹,成虫の4段階に区分される.生活史の各段階(発生ステージ)の長さは,温度や日長などの棲息する環境に影響を受ける.
※4 一塩基多型(SNPs): 生物のもつDNAの塩基配列が一塩基単位で変異した違い.この変異はゲノム中に多く存在し,とても近縁な遺伝的関係(親子関係)の間にも違いがありことが知られている.一塩基多型(Single Nucleotide Polymorphism; SNPs)の変異を調べることで,ミトコンドリアDNAの塩基配列では区別することができなかった,近接する集団同士の遺伝的な違いの検出も可能となっている.
※5 ゲノムワイド関連解析(Genome-Wide Association Study, GWAS): 集団内にある表現型の多型とゲノム全域から得られた遺伝的多型との間の相関関係を調べ,形質に関連する遺伝子を究明する解析.
【備考】
・高山帯に関する調査は環境省許可番号 環中中国許2109012, 2206271のもと実施.
・サハリントビケラの分布域を網羅した分子系統・生物地理学的研究については,論文として公開済みである.研究結果については以下を参照:Suzuki, H., Takenaka, M., & Tojo, K. (2024) Evolutionary history of a cold-adapted limnephilid caddisfly: Effects of climate change and topography on genetic structure. Molecular Phylogenetics and Evolution, 191: 107967.
https://doi.org/10.1016/j.ympev.2023.107967
亜高山帯における生活史調査の結果,亜高山帯では成虫が盛夏に「夏眠(卵巣休眠)」している可能性が示唆された.「夏眠」とは,夏季において高温や乾燥に耐えるように代謝を一時的に低下(もしくは停止)させる休眠パターンであり,サハリントビケラを含む近縁なトビケラ類においても成虫期に夏眠し,卵巣発達を一時的に停止させる,いわゆる「卵巣休眠」を行うことが知られている.サハリントビケラの生活史について追究した先行研究では,初夏に出現した成虫は,盛夏に出現頻度は極めて減少し,再び秋に出現頻度が増加するとしている.北米に棲息する同属別種においても,同様の傾向が知られている.これらのことから,夏眠を有する生活史はサハリントビケラの一般的な生活史パターンであると推察される.夏眠を有する生活史パターンは,従来,寒冷な環境に棲息していたサハリントビケラが,高温や乾燥にさらされる可能性が高い地域に分布域を拡大し,環境に適応した結果,獲得した形質と考えられる.
一方,高山帯は亜高山帯とは異なり,成虫が集中的に出現し,卵未成熟の成虫が出現する期間は亜高山帯と比べて短期間であった.従って高山帯の集団には,夏眠期間が「ない」もしくは「短縮」されている可能性が示された.本研究によって示唆された,高山帯に棲息する集団の夏眠の欠如(もしくは短縮)する生活史パターンは,活動に適する期間が短い地域で繁殖を成功させる可能性の高い生態的特徴であると推察される.ノルウェーなど北極圏に近い地域に分布するサハリントビケラの近縁種の中には夏眠を行わない種があることも知られている.高緯度地域,高山帯はいずれも寒冷で夏が短い.こうしたことから,気温や日長などの要因が複合的に機能し,成虫の夏眠の有無に寄与していると考えられる.
以上より,サハリントビケラが棲息環境に応じて異なる適応をしていることが示唆された.次に,生活史の違いと標高帯間の遺伝的分化との関連性について追究した.
生息環境と夏眠(卵巣休眠)の有無
サハリントビケラは,オスが精子の入った袋「精包」をメスへと渡すことで交尾が完了する.つまり,メスの腹部にある精包の有無を調べることで,交尾時期を知ることができる.それぞれの標高帯で精包の有無を確認したところ,いずれの標高帯においても,成虫が観察され始めた直後(高山帯:7月下旬,亜高山帯:6月中旬)に,卵が未成熟であるメスの体内から精包が確認された(図4).このことは,本種は羽化直後の状態で,既に交尾を済ませており,また,それぞれの標高帯では交尾時期が異なる(ズレている)ことも示している.また,夏眠の有無の違いにより,成虫の活動時期も標高帯間で分化していることから,それぞれの集団間で交配が生じる可能性は低い.したがって,集団間の交配前生殖隔離が働いており,集団間の遺伝的分化が維持・促進されていると推測される.現在,標高帯間に存在する遺伝子流動の程度(遺伝的障壁の大きさ)を評価するべく,ゲノムワイドに存在する一塩基多型(SNPs)※4情報に基づく遺伝子解析を進めている.
結論
本研究は,同一山塊内の高山帯・亜高山帯という極めて近接する集団を形成する水生昆虫を対象に,生活史の差異(ズレ)を明示し,それぞれの生活史が異なる環境下において適応的な意義をもつ可能性を示した初めての事例である.今後,より詳細な遺伝子解析の結果を加味し,標高帯間に存在する遺伝的障壁を明らかにしていく予定である.
将来的な研究として,「生活史の差異は遺伝的に固定化されているのか?」,また「それぞれの環境への適応に関与する遺伝子は存在するのか?」といったような疑問を追究すべく,飼育実験やゲノムワイド関連解析(GEAS)※5に発展させ,山岳における生物多様性創出メカニズムの解明に前進していくことを目指す.
【用語解説】
※1 トビケラ: 毛翅目の昆虫類(鱗翅目の蝶や蛾に近縁な昆虫のグループ)で,幼虫は芋虫型,ほとんどの種が水中で生活する.口から吐く糸で筒状の巣(筒巣)を形成し,巣内で生活する種も多い.成虫は陸上生活するが,翅には(チョウ類やガ類がもつような)鱗粉ではなく,細かな毛が生えている.
※2 化性: 昆虫が1年間に世代を繰り返す性質のこと.1年間に1世代が繁殖し,一年周期で世代が繋がるような生活史をもつことを年1化(あるいは,単に「年一世代」)という.
※3 生活史: 生物が生まれてから死ぬまでの過程.昆虫の場合,卵,幼虫,蛹,成虫の4段階に区分される.生活史の各段階(発生ステージ)の長さは,温度や日長などの棲息する環境に影響を受ける.
※4 一塩基多型(SNPs): 生物のもつDNAの塩基配列が一塩基単位で変異した違い.この変異はゲノム中に多く存在し,とても近縁な遺伝的関係(親子関係)の間にも違いがありことが知られている.一塩基多型(Single Nucleotide Polymorphism; SNPs)の変異を調べることで,ミトコンドリアDNAの塩基配列では区別することができなかった,近接する集団同士の遺伝的な違いの検出も可能となっている.
※5 ゲノムワイド関連解析(Genome-Wide Association Study, GWAS): 集団内にある表現型の多型とゲノム全域から得られた遺伝的多型との間の相関関係を調べ,形質に関連する遺伝子を究明する解析.
【備考】
・高山帯に関する調査は環境省許可番号 環中中国許2109012, 2206271のもと実施.
・サハリントビケラの分布域を網羅した分子系統・生物地理学的研究については,論文として公開済みである.研究結果については以下を参照:Suzuki, H., Takenaka, M., & Tojo, K. (2024) Evolutionary history of a cold-adapted limnephilid caddisfly: Effects of climate change and topography on genetic structure. Molecular Phylogenetics and Evolution, 191: 107967.
https://doi.org/10.1016/j.ympev.2023.107967
【論文タイトルと著者等】
タイトル: Variations in the phenological patterns of a caddisfly inhabiting the same mountain massifs: life history differences in different altitudinal zones
著者: Hirohisa SUZUKI, Masaki TAKENAKA, Koji TOJO
掲載誌: Ecology and Evolution 191: 107967
掲載日: 2024年6月6日公開
URL: https://onlinelibrary.wiley.com/doi/10.1002/ece3.11428
D O I: doi.org/10.1002/ece3.11428
Hirohisa SUZUKI 鈴木啓久(岐阜県立飛騨高山高校 教諭)筆頭著者
Masaki TAKENAKA 竹中將起(信州大学 特任助教)
Koji TOJO 東城幸治(信州大学 教授)責任著者
タイトル: Variations in the phenological patterns of a caddisfly inhabiting the same mountain massifs: life history differences in different altitudinal zones
著者: Hirohisa SUZUKI, Masaki TAKENAKA, Koji TOJO
掲載誌: Ecology and Evolution 191: 107967
掲載日: 2024年6月6日公開
URL: https://onlinelibrary.wiley.com/doi/10.1002/ece3.11428
D O I: doi.org/10.1002/ece3.11428
Hirohisa SUZUKI 鈴木啓久(岐阜県立飛騨高山高校 教諭)筆頭著者
Masaki TAKENAKA 竹中將起(信州大学 特任助教)
Koji TOJO 東城幸治(信州大学 教授)責任著者