生物学コース 東城幸治教授らの研究グループが、日本列島のサワガニの分布域を網羅する遺伝子解析により、系統進化史を解明しました。
2023年10月10日
生涯を渓流で過ごすサワガニなのに,「海流分散」の歴史あり!
陸・海の2つのルートで分布拡大,そして新天地での2次的接触複数の未記載種(新種)を新たに発見!
【研究成果のポイント】
・サワガニ(沢蟹)は淡水に生息する甲殻類で、日本列島広域に生息(北海道には非分布)
・日本列島のサワガニの分布域を網羅するような遺伝子解析により、系統進化史を解明
・サワガニの祖先は琉球列島などの南西地域に起源し、北方へと分布拡大(分散)
・サワガニにとって海峡は大きな分散障壁として機能するため、陸橋を介した分散が基本
・ただし、陸路分散だけでは理解不可能な遺伝構造を検出 → 海流(黒潮)分散の可能性
・サワガニの海水(塩分)耐性を実験的に確認 → 海流分散に耐えられる潜在能力あり
・質の異なる「陸」・「海」2ルートでの分散という珍しい現象を確認
・新たに未記載種(新種,隠蔽種)を発見 → 今後の分類学的な整理が必要
・サワガニ(沢蟹)は淡水に生息する甲殻類で、日本列島広域に生息(北海道には非分布)
・日本列島のサワガニの分布域を網羅するような遺伝子解析により、系統進化史を解明
・サワガニの祖先は琉球列島などの南西地域に起源し、北方へと分布拡大(分散)
・サワガニにとって海峡は大きな分散障壁として機能するため、陸橋を介した分散が基本
・ただし、陸路分散だけでは理解不可能な遺伝構造を検出 → 海流(黒潮)分散の可能性
・サワガニの海水(塩分)耐性を実験的に確認 → 海流分散に耐えられる潜在能力あり
・質の異なる「陸」・「海」2ルートでの分散という珍しい現象を確認
・新たに未記載種(新種,隠蔽種)を発見 → 今後の分類学的な整理が必要

【背景】
日本列島の広域に生息するサワガニは,最も「身近な生きもの」の代表格であり,しばしば「昔ばなし」にも登場する(北海道には分布しないとされるが,近年,確認されている:人為分布の可能性あり)。河川の源流域や渓流のほか,里山の水路や水田などでもよく見かけるお馴染みの動物であるが,その進化史は,実に複雑なものであることが,今回発表された論文において明らかとなった。
日本産サワガニ類(中琉球・南琉球地域を除く)
サワガニ属 Geothelphusa Stimpson, 1858 日本列島(琉球列島を含む),台湾
1)サワガニ Geothelphusa dehaani (White, 1847) 本州,四国,九州,周辺離島
(屋久島,種子島,中之島,甑島,隠岐島, 福江島,佐渡島など)
2)ミカゲサワガニ Geothelphusa exigua Suzuki and Tsuda, 1994 大隅半島
3)ヤクシマサワガニ Geothelphusa marmorata Suzuki and Okano, 2000 屋久島
4)コシキサワガニ Geothelphusa koshikiensis Suzuki and Kawai, 2011 甑島
5)ミシマサワガニ Geothelphusa mishima Suzuki and Kawai, 2011 黒島ほか
このうち,最も広域に生息する1)のサワガニには,図1に示すように様々な体色のものが知られている。例えば,屋久島や中之島(トカラ列島),甑島(こしきじま)列島の集団は青白い体色をしているが,九州本土の集団では赤色であるなど,体色は大きく異なっている。また,四国地方や本州東部では,同一地点において複数の体色の個体が混生する事例も報告されており,体色と遺伝的な関係性については否定されてきた。
このような背景下,日本列島のサワガニの分布域を網羅するような広域的な地域集団を対象にサンプリングを実施し,合計126地点(地域集団)の268個体を試料として用い,遺伝子を抽出・精製し,分子系統解析を実施した。
日本列島の広域に生息するサワガニは,最も「身近な生きもの」の代表格であり,しばしば「昔ばなし」にも登場する(北海道には分布しないとされるが,近年,確認されている:人為分布の可能性あり)。河川の源流域や渓流のほか,里山の水路や水田などでもよく見かけるお馴染みの動物であるが,その進化史は,実に複雑なものであることが,今回発表された論文において明らかとなった。
日本産サワガニ類(中琉球・南琉球地域を除く)
サワガニ属 Geothelphusa Stimpson, 1858 日本列島(琉球列島を含む),台湾
1)サワガニ Geothelphusa dehaani (White, 1847) 本州,四国,九州,周辺離島
(屋久島,種子島,中之島,甑島,隠岐島, 福江島,佐渡島など)
2)ミカゲサワガニ Geothelphusa exigua Suzuki and Tsuda, 1994 大隅半島
3)ヤクシマサワガニ Geothelphusa marmorata Suzuki and Okano, 2000 屋久島
4)コシキサワガニ Geothelphusa koshikiensis Suzuki and Kawai, 2011 甑島
5)ミシマサワガニ Geothelphusa mishima Suzuki and Kawai, 2011 黒島ほか
このうち,最も広域に生息する1)のサワガニには,図1に示すように様々な体色のものが知られている。例えば,屋久島や中之島(トカラ列島),甑島(こしきじま)列島の集団は青白い体色をしているが,九州本土の集団では赤色であるなど,体色は大きく異なっている。また,四国地方や本州東部では,同一地点において複数の体色の個体が混生する事例も報告されており,体色と遺伝的な関係性については否定されてきた。
このような背景下,日本列島のサワガニの分布域を網羅するような広域的な地域集団を対象にサンプリングを実施し,合計126地点(地域集団)の268個体を試料として用い,遺伝子を抽出・精製し,分子系統解析を実施した。

図1.日本列島におけるサワガニの体色多型.地域集団によって体色は多様.A) 長野県,B) 佐渡島,C) 九州,D) 伊豆半島の地域集団
【結果・考察】
サワガニ類における分子系統解析
ミトコンドリアDNA(mtDNA)COI領域,16S rRNA領域,および核DNA(nDNA) histone H3領域の塩基配列(1,796-bp)に基づく系統解析を実施した結果,サワガニの近縁種(同属別種)が国内では南西諸島に比較的種類が多く,また海外のサワガニ類についても東南アジア地域の種多様性が高いことから,日本産サワガニ類の祖先系統は,列島の南西地域に起源し,より北方へと分布域を拡大していった進化プロセスが支持された(図2)。
サワガニ類における分子系統解析
ミトコンドリアDNA(mtDNA)COI領域,16S rRNA領域,および核DNA(nDNA) histone H3領域の塩基配列(1,796-bp)に基づく系統解析を実施した結果,サワガニの近縁種(同属別種)が国内では南西諸島に比較的種類が多く,また海外のサワガニ類についても東南アジア地域の種多様性が高いことから,日本産サワガニ類の祖先系統は,列島の南西地域に起源し,より北方へと分布域を拡大していった進化プロセスが支持された(図2)。
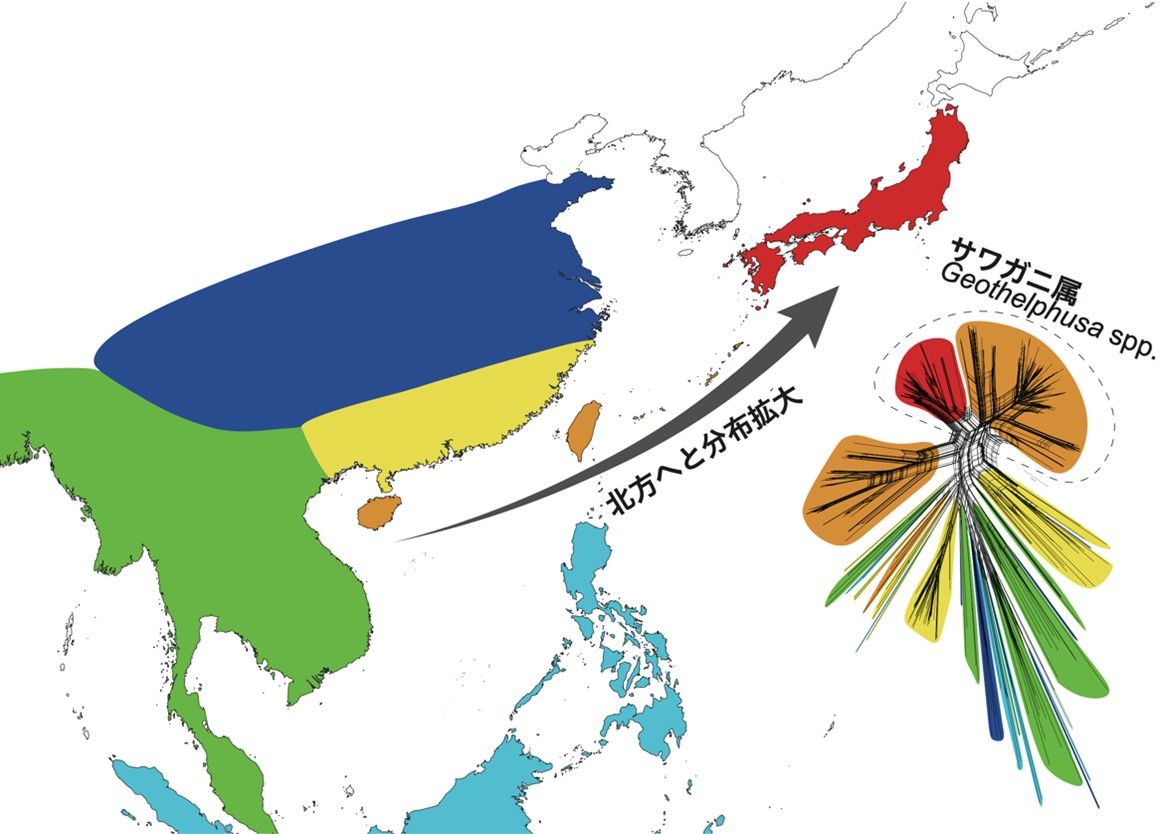
図2.アジア地域のサワガニ類の分布と系統関係.東南アジアから東アジアにかけての系統進化史としては種多様性の高い南方に起源し,種分化しながら分布域を北上させてきた(本州はサワガニ類の分布北限に相当する)
また本研究では,サワガニG. dehaani 種内に 10の主要な遺伝系統が検出され,いずれも互いに大きく遺伝分化した系統であることが明らかとなった(図3)。基本的には,本州,四国,九州といった主要島ごとの遺伝分化に加えて,より小さな離島にも固有の系統が検出された(一部,本州や四国,九州の島を跨いで検出される系統もみられた)。このような遺伝的に大きく分化した系統が検出された理由としては,飛翔能力をもたないサワガニの移動能力の低さがあらわれたものと考えられる。一方,陸続きとなる同一島内での遺伝的分化はみられず,陸続きであれば移動分散が可能であることが示唆された。すなわち,基本的には海峡が遺伝子流動の障壁として機能していることが明示された。サワガニは陸上を歩行することができるため,陸続きで森林面積が大きな島では,広域的に移動可能であることを示している。
また,分岐年代を推定した結果,それぞれの系統が分岐した年代は,瀬戸内海が形成される以前であったと推定された。ちょうど日本列島の地殻変動が活発であった時期であり,日本列島西部において山岳形成が活発な時期に相当するため,他の多くの生物と同様に山岳形成に伴い各地域ごとに遺伝分化したと推定された。
しかし,例外的な系統(系統4)もみられ,これは伊豆半島,三浦半島,房総半島の集団から検出されたものであるが,この系統4に最も近縁であるとして評価されたのは,(地理的には遠く離れた)九州や北琉球の集団で構成される系統3と評価された(図3)。こうした近縁な遺伝系統群が「飛び地」的に分布し,かつ太平洋側へと突き出た半島部にみられることは興味深く,黒潮による海流分散によるものと示唆された。
また,分岐年代を推定した結果,それぞれの系統が分岐した年代は,瀬戸内海が形成される以前であったと推定された。ちょうど日本列島の地殻変動が活発であった時期であり,日本列島西部において山岳形成が活発な時期に相当するため,他の多くの生物と同様に山岳形成に伴い各地域ごとに遺伝分化したと推定された。
しかし,例外的な系統(系統4)もみられ,これは伊豆半島,三浦半島,房総半島の集団から検出されたものであるが,この系統4に最も近縁であるとして評価されたのは,(地理的には遠く離れた)九州や北琉球の集団で構成される系統3と評価された(図3)。こうした近縁な遺伝系統群が「飛び地」的に分布し,かつ太平洋側へと突き出た半島部にみられることは興味深く,黒潮による海流分散によるものと示唆された。
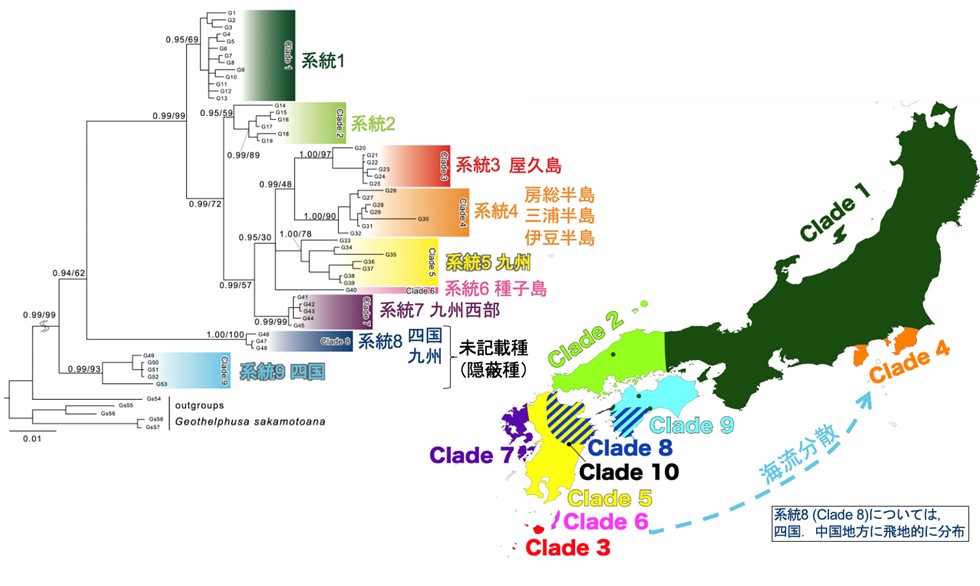
図3.日本列島内のサワガニ Geothelphusa dehaani の分子系統解析の結果.ミトコンドリアDNA(mtDNA)COI領域,16S rRNA領域,および核DNA( nDNA) histone H3領域の塩基配列(1,796-bp)に基づく系統解析の結果(ベイズ系統樹)と,各遺伝系統群(Clade)を構成するサワガニが採取された地域を示し,系統樹と地図の配色は対応している.系統4を構成する房総半島・三浦半島・伊豆半島のサワガニは屋久島の集団で構成される系統3に最も近縁であり,次いで近いのが系統5(九州広域)や系統6(種子島)となり,系統4だけが飛び地的な分布となる.しかし,黒潮に漂流して分散したと考えれば理解は容易となる.
サワガニ類における塩分(海水)耐性実験
海流の流路に沿って離れた地域間で共通性の高い遺伝構造をとるような生物には,プランクトン生活を有するものがよく知られている。カニ類が属する十脚類(いわゆるエビ・カニ類)の多くは,プランクトン(浮遊)生活をする幼生期を有する海産種であるが,今回の研究対象であるサワガニ類は,一生を淡水域で暮らす純淡水種である。また,一部の文献や図鑑には「海水では生きていけない」とも記述されているため,実際に海水中での生存の可否を,耐塩実験により検証してみた。
その結果,サワガニは海水と同程度の塩分濃度ではほぼ問題なく生存できることが究明された(実施した2週間の実験では高い生存率が示された)。この実験には,海流分散が示唆される遺伝系統群だけでなく,陸路で分散してきたと考えられる集団も対象としたが,いずれにおいても高い塩分耐性が示された。これらのことから,サワガニ全体が潜在的に高い塩分耐性をもち,こうした特性が海流分散を可能にしたと推察される。
想定される黒潮による海流分散
遺伝子解析の結果,伊豆半島,三浦半島,房総半島のサワガニは,北琉球(屋久島,吐?喇列島の口之島や中之島)のサワガニとの近縁性が示唆された。さらに,次に近縁な集団も九州本土や種子島であり,海流による分散を想定しなければ説明し難い結果と言える。南西諸島の島嶼や半島部では,渓流のような景観のまま海に注ぐような小規模河川も多く,こうした小規模渓流の海岸線付近にもサワガニは多く生息している。こうした生息地に起こる洪水では,河畔を構成する樹木の折れ枝などと共に,サワガニも海へと流されてしまうことも頻繁に生じているものと想像される。2週間もの期間を海水中で生存することが可能であるならば,長い時間スケールの中では,黒潮の流路先へと漂流する可能性も考えられる。
また,海流分散が可能であることと,漂流先に定着できるかどうかは別課題である。陸橋を介した陸路での分散においても,本州最北まで分布拡大してきた背景があるため,先に陸路で分散したサワガニが分布しているとすれば,そこへ海流によりわずかに漂着した個体が定着し,遺伝子頻度をかえることは容易ではない。ただし,伊豆半島,三浦半島,房総半島は,かつて独立した島であったことから,空いたニッチを利用できた可能性も考えられる。北琉球の系統や,海流分散により分布拡大した系統では,青白い体色のサワガニが優占しており,系統的な形質である可能性や,創始者効果,海流分散に関わる何かしらの意義も考え得るが,体色に関しては今後の課題である。また,陸路と海路それぞれで分散した系統が二次的に接触する地域では,体色に2型がみられるような集団も存在する。今後,体色の違いと系統の関係性を調査することは興味深い課題である。遺伝子流動も含め,核遺伝子の解析やゲノムワイドな遺伝マーカーを用いた,より詳細な解析への展開が求められる。
分類学的な再検討・未記載種(新種)の発見
最後に,本研究から複数の未記載種(いわゆる新種,隠蔽種)が発見された(図3)。従来,サワガニG. dehaaniと扱われてきた種内において,別種レベルでの遺伝分化にも匹敵する遺伝分化が検出された。琉球列島内の島嶼と同様に,本州,四国,九州においても島ごとに種分化している可能性も示唆され,四国や九州には複数の未記載種,もしくは隠蔽種が存在している可能性は高い。サワガニ類では,オスの第一腹節(付属肢)first gonopodsの形態形質が分類学上に鍵形質として種識別がされているため,今後はこうした形質にも注目して分類学的な再検討を進めるとともに,今回明らかとなった遺伝系統間での交配が生じているのかどうかについても解析を深めていく予定である。
【用語解説】
隠蔽種:本来は別の種であるものの,形態による区別がつかないため,同一種として扱われてきた種を指す。 遺伝子マーカーを用いた分子系統学的な研究により,隠蔽種であることが判明する事例が多い。
海流の流路に沿って離れた地域間で共通性の高い遺伝構造をとるような生物には,プランクトン生活を有するものがよく知られている。カニ類が属する十脚類(いわゆるエビ・カニ類)の多くは,プランクトン(浮遊)生活をする幼生期を有する海産種であるが,今回の研究対象であるサワガニ類は,一生を淡水域で暮らす純淡水種である。また,一部の文献や図鑑には「海水では生きていけない」とも記述されているため,実際に海水中での生存の可否を,耐塩実験により検証してみた。
その結果,サワガニは海水と同程度の塩分濃度ではほぼ問題なく生存できることが究明された(実施した2週間の実験では高い生存率が示された)。この実験には,海流分散が示唆される遺伝系統群だけでなく,陸路で分散してきたと考えられる集団も対象としたが,いずれにおいても高い塩分耐性が示された。これらのことから,サワガニ全体が潜在的に高い塩分耐性をもち,こうした特性が海流分散を可能にしたと推察される。
想定される黒潮による海流分散
遺伝子解析の結果,伊豆半島,三浦半島,房総半島のサワガニは,北琉球(屋久島,吐?喇列島の口之島や中之島)のサワガニとの近縁性が示唆された。さらに,次に近縁な集団も九州本土や種子島であり,海流による分散を想定しなければ説明し難い結果と言える。南西諸島の島嶼や半島部では,渓流のような景観のまま海に注ぐような小規模河川も多く,こうした小規模渓流の海岸線付近にもサワガニは多く生息している。こうした生息地に起こる洪水では,河畔を構成する樹木の折れ枝などと共に,サワガニも海へと流されてしまうことも頻繁に生じているものと想像される。2週間もの期間を海水中で生存することが可能であるならば,長い時間スケールの中では,黒潮の流路先へと漂流する可能性も考えられる。
また,海流分散が可能であることと,漂流先に定着できるかどうかは別課題である。陸橋を介した陸路での分散においても,本州最北まで分布拡大してきた背景があるため,先に陸路で分散したサワガニが分布しているとすれば,そこへ海流によりわずかに漂着した個体が定着し,遺伝子頻度をかえることは容易ではない。ただし,伊豆半島,三浦半島,房総半島は,かつて独立した島であったことから,空いたニッチを利用できた可能性も考えられる。北琉球の系統や,海流分散により分布拡大した系統では,青白い体色のサワガニが優占しており,系統的な形質である可能性や,創始者効果,海流分散に関わる何かしらの意義も考え得るが,体色に関しては今後の課題である。また,陸路と海路それぞれで分散した系統が二次的に接触する地域では,体色に2型がみられるような集団も存在する。今後,体色の違いと系統の関係性を調査することは興味深い課題である。遺伝子流動も含め,核遺伝子の解析やゲノムワイドな遺伝マーカーを用いた,より詳細な解析への展開が求められる。
分類学的な再検討・未記載種(新種)の発見
最後に,本研究から複数の未記載種(いわゆる新種,隠蔽種)が発見された(図3)。従来,サワガニG. dehaaniと扱われてきた種内において,別種レベルでの遺伝分化にも匹敵する遺伝分化が検出された。琉球列島内の島嶼と同様に,本州,四国,九州においても島ごとに種分化している可能性も示唆され,四国や九州には複数の未記載種,もしくは隠蔽種が存在している可能性は高い。サワガニ類では,オスの第一腹節(付属肢)first gonopodsの形態形質が分類学上に鍵形質として種識別がされているため,今後はこうした形質にも注目して分類学的な再検討を進めるとともに,今回明らかとなった遺伝系統間での交配が生じているのかどうかについても解析を深めていく予定である。
【用語解説】
隠蔽種:本来は別の種であるものの,形態による区別がつかないため,同一種として扱われてきた種を指す。 遺伝子マーカーを用いた分子系統学的な研究により,隠蔽種であることが判明する事例が多い。
【論文タイトルと著者等】
タイトル:Phylogeography of the true freshwater crab, Geothelphusa dehaani: detected dual dispersal routes via land and sea
著者: Masaki TAKENAKA, Koki YANO, Koji TOJO
掲載誌:Zoology 160: 126118
掲載日:2023年8月23日
URL:https://doi.org/10.1016/j.zool.2023.126118
D O I:10.1016/j.zool.2023.126118
Masaki TAKENAKA 竹中將起(信州大学,筑波大学) 筆頭著者
Koki YANO 谷野宏樹(基礎生物学研究所)筆頭著者
Koji TOJO 東城幸治(信州大学)責任著者
【問い合わせ先】
〈研究内容に関する問い合わせ先〉
信州大学学術研究院理学系(理学部理学科生物学コース)
特任助教 竹中將起 Tel:0263-37-2493,E-mail:takenaka10mt@shinshu-u.ac.jp
教授 東城幸治 Tel:0263-37-3341,E-mail:ktojo@shinshu-u.ac.jp
生物学コース事務室 Tel:0263-37-2488
〈報道に関する問い合わせ先〉
国立大学法人信州大学 総務部総務課広報室
〒390-8621 長野県松本市旭3-1-1
Tel:0263-37-3056 Fax:0263-37-2182 E-mail:shinhp@shinshu-u.ac.jp
国立大学法人筑波大学広報局
TEL:029-853-2040 Fax:029-853-2014 E-mail:kohositu@un.tsukuba.ac.jp
自然科学研究機構 基礎生物学研究所 広報室
TEL:0564-55-7628 Fax:0564-55-7597 E-mail:press@nibb.ac.jp
タイトル:Phylogeography of the true freshwater crab, Geothelphusa dehaani: detected dual dispersal routes via land and sea
著者: Masaki TAKENAKA, Koki YANO, Koji TOJO
掲載誌:Zoology 160: 126118
掲載日:2023年8月23日
URL:https://doi.org/10.1016/j.zool.2023.126118
D O I:10.1016/j.zool.2023.126118
Masaki TAKENAKA 竹中將起(信州大学,筑波大学) 筆頭著者
Koki YANO 谷野宏樹(基礎生物学研究所)筆頭著者
Koji TOJO 東城幸治(信州大学)責任著者
【問い合わせ先】
〈研究内容に関する問い合わせ先〉
信州大学学術研究院理学系(理学部理学科生物学コース)
特任助教 竹中將起 Tel:0263-37-2493,E-mail:takenaka10mt@shinshu-u.ac.jp
教授 東城幸治 Tel:0263-37-3341,E-mail:ktojo@shinshu-u.ac.jp
生物学コース事務室 Tel:0263-37-2488
〈報道に関する問い合わせ先〉
国立大学法人信州大学 総務部総務課広報室
〒390-8621 長野県松本市旭3-1-1
Tel:0263-37-3056 Fax:0263-37-2182 E-mail:shinhp@shinshu-u.ac.jp
国立大学法人筑波大学広報局
TEL:029-853-2040 Fax:029-853-2014 E-mail:kohositu@un.tsukuba.ac.jp
自然科学研究機構 基礎生物学研究所 広報室
TEL:0564-55-7628 Fax:0564-55-7597 E-mail:press@nibb.ac.jp