生物学コース 東城幸治教授らの研究グループが、高山帯・亜高山帯に適応した昆虫(サハリントビケラ)の進化史をDNA解析で究明しました。
2024年1月26日
「山岳」は生物多様性を生み出す「泉」! 「気候変動」が種多様性を生み出す!
―高山帯・亜高山帯に適応した昆虫(サハリントビケラ)の進化史をDNA解析で究明: 山岳特有の「地形」と「水環境」が遺伝的多様化に深く関与―
【研究成果のポイント】
・サハリントビケラは冷涼な池沼や泉に生息する水生昆虫で,中部山岳~サハリンに分布
・種内に別種レベルの遺伝的分化を検出(系統I・系統II)
・サハリントビケラの分布域シフトには気候変動(氷期?間氷期サイクル)が影響
・系統Iは北海道・東北で遺伝的多様化,系統IIは中部山岳域で遺伝的多様化
・系統間での遺伝構造の差異は生息場の標高差による(特に,高山帯では強い遺伝的隔離)
・系統IIの遺伝構造から,氷期にも山塊内の不凍域(湧水など)で集団を維持していた可能性あり
・気候変動は,高山帯における種内系統間でのニッチ分化・遺伝分化をもたらす
・サハリントビケラは冷涼な池沼や泉に生息する水生昆虫で,中部山岳~サハリンに分布
・種内に別種レベルの遺伝的分化を検出(系統I・系統II)
・サハリントビケラの分布域シフトには気候変動(氷期?間氷期サイクル)が影響
・系統Iは北海道・東北で遺伝的多様化,系統IIは中部山岳域で遺伝的多様化
・系統間での遺伝構造の差異は生息場の標高差による(特に,高山帯では強い遺伝的隔離)
・系統IIの遺伝構造から,氷期にも山塊内の不凍域(湧水など)で集団を維持していた可能性あり
・気候変動は,高山帯における種内系統間でのニッチ分化・遺伝分化をもたらす
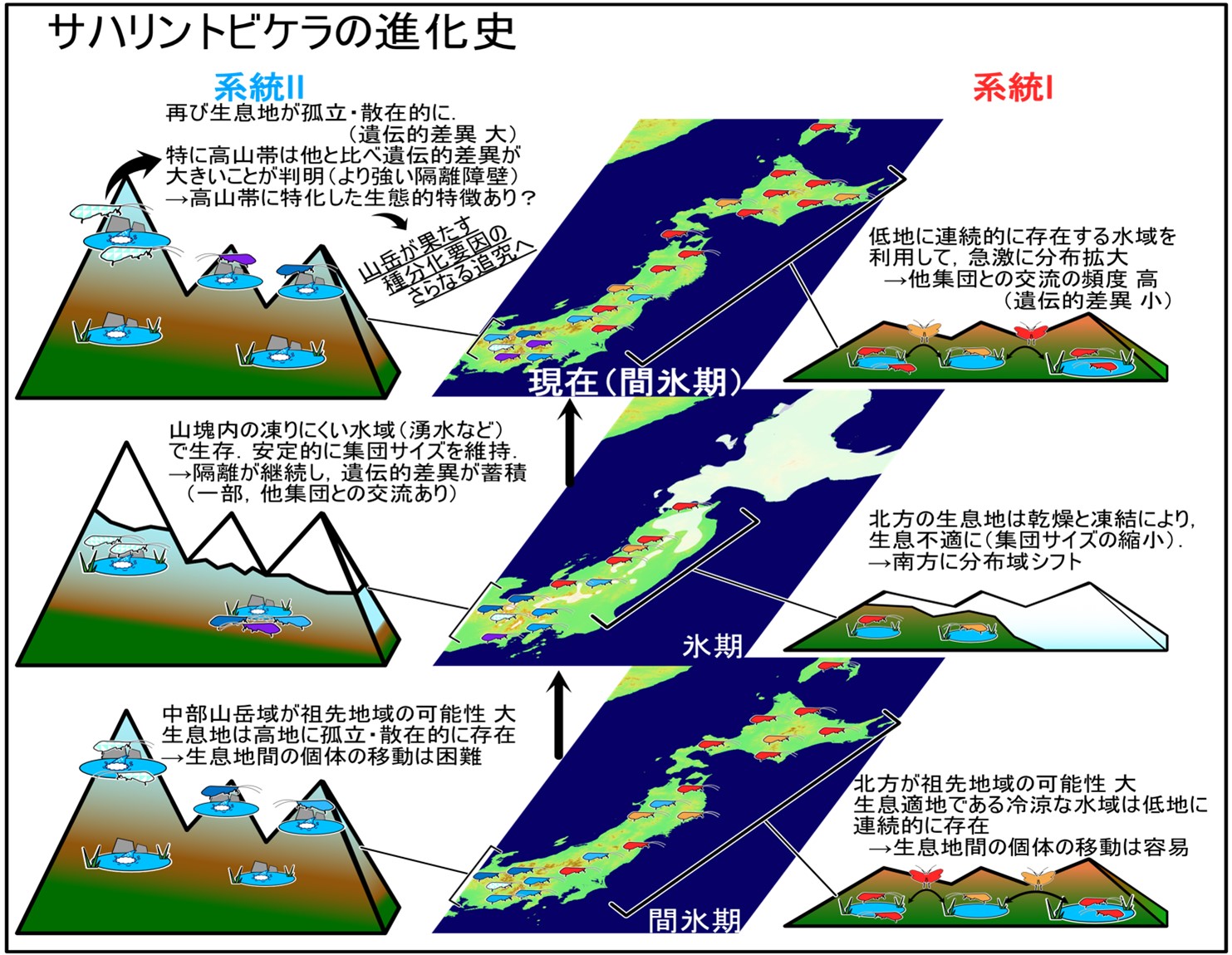
【背景】
生きものたちの棲み場所としての「高山帯」は,低温,強風,冬季の凍結,乾燥,強い紫外線などに晒される極めて過酷な環境である.高山植物やニホンライチョウなどのように,過酷な環境に適応した,ごく限られた生物種だけが棲む特殊なハビタット*1と考えられてきた.こうした高山帯は,日本アルプスのように規模の大きな山脈であっても山域ごとにパッチ状に位置することから,しばしば「Sky Islands(天空に浮かぶ島々)」などと喩えられる.このような過酷な環境下で生存できる生物種は限られるため,種の固有性や独立性は高いものの,種を生み出す(種多様性を創生する)場としては注目されてこなかった.
こうした背景下,この研究では,高山帯とそれに準ずる亜高山帯の冷涼な環境に適応した昆虫に注目し,山岳が有するダイナミックな地形的特徴と,過去の長期的な気候変動(氷期-間氷期サイクル)の相互作用が,生物多様性を生み出す場にもなり得ることを示した.
本研究では,日本列島の中部山岳域からサハリンにまで分布する寒冷適応水生昆虫の一種 サハリントビケラを対象に,この種の分布域を網羅するような地域集団を対象とした遺伝子解析を実施し,種内の系統進化史を詳細に紐解くとともに,気候変動がサハリントビケラの分布域のシフトに与えた影響を考察した.さらに,高山帯から亜高山帯に生息する水生生物の遺伝的多様化には,陸上生物とは異なる(氷期にも不凍である「湧水」をリフュージア*2として利用することで,生存不適な期間を生き延びる)メカニズムが働いた可能性を示唆した.また,高山帯に生息する生物種における,集団レベルでの遺伝構造は,山岳に生息する系統の中でも特殊であり,強い遺伝的隔離が働いている可能性が示唆された.すなわち,高山帯で引き起こされる「種分化」の要因解明にも寄与する成果といえる.
【研究対象種:サハリントビケラ】
サハリントビケラAsynarchus sachalinensis Martynov, 1914は,渓流魚釣りのエサとして馴染みのある「トビケラ*3」の一種であり,冷涼な池沼や泉に生息する(図1).そして,本州中部にある高山帯の池沼にも生息するため,きわめて寒冷な環境下でも生存することができる.
今回,分布域である本州の中部山岳からサハリンを網羅するように広域的なサンプリングを実施し,合計74地点(地域集団)から採取した144個体を解析試料として用い,全ゲノムDNAを抽出・精製し,分子系統解析・遺伝構造解析を実施した.
生きものたちの棲み場所としての「高山帯」は,低温,強風,冬季の凍結,乾燥,強い紫外線などに晒される極めて過酷な環境である.高山植物やニホンライチョウなどのように,過酷な環境に適応した,ごく限られた生物種だけが棲む特殊なハビタット*1と考えられてきた.こうした高山帯は,日本アルプスのように規模の大きな山脈であっても山域ごとにパッチ状に位置することから,しばしば「Sky Islands(天空に浮かぶ島々)」などと喩えられる.このような過酷な環境下で生存できる生物種は限られるため,種の固有性や独立性は高いものの,種を生み出す(種多様性を創生する)場としては注目されてこなかった.
こうした背景下,この研究では,高山帯とそれに準ずる亜高山帯の冷涼な環境に適応した昆虫に注目し,山岳が有するダイナミックな地形的特徴と,過去の長期的な気候変動(氷期-間氷期サイクル)の相互作用が,生物多様性を生み出す場にもなり得ることを示した.
本研究では,日本列島の中部山岳域からサハリンにまで分布する寒冷適応水生昆虫の一種 サハリントビケラを対象に,この種の分布域を網羅するような地域集団を対象とした遺伝子解析を実施し,種内の系統進化史を詳細に紐解くとともに,気候変動がサハリントビケラの分布域のシフトに与えた影響を考察した.さらに,高山帯から亜高山帯に生息する水生生物の遺伝的多様化には,陸上生物とは異なる(氷期にも不凍である「湧水」をリフュージア*2として利用することで,生存不適な期間を生き延びる)メカニズムが働いた可能性を示唆した.また,高山帯に生息する生物種における,集団レベルでの遺伝構造は,山岳に生息する系統の中でも特殊であり,強い遺伝的隔離が働いている可能性が示唆された.すなわち,高山帯で引き起こされる「種分化」の要因解明にも寄与する成果といえる.
【研究対象種:サハリントビケラ】
サハリントビケラAsynarchus sachalinensis Martynov, 1914は,渓流魚釣りのエサとして馴染みのある「トビケラ*3」の一種であり,冷涼な池沼や泉に生息する(図1).そして,本州中部にある高山帯の池沼にも生息するため,きわめて寒冷な環境下でも生存することができる.
今回,分布域である本州の中部山岳からサハリンを網羅するように広域的なサンプリングを実施し,合計74地点(地域集団)から採取した144個体を解析試料として用い,全ゲノムDNAを抽出・精製し,分子系統解析・遺伝構造解析を実施した.

図1.サハリントビケラの典型的な生息地と成虫・幼虫の写真.(a)亜高山帯にある系統Iの生息地(長野県:乗鞍高原 標高 約1,200 m),(b)高山帯にある系統IIの生息地(岐阜県:乗鞍岳 標高 約2,700 m),(c)オス,(d)メス,(e)幼虫とその筒巣
【結果・考察】
サハリントビケラにおける分子系統解析
ミトコンドリアDNA(mtDNA)COI領域(648-bp),16S rRNA領域(417-bp)を結合した塩基配列に基づく系統解析を実施した結果,サハリントビケラの種内には別種として扱われてもよいレベルに大きく遺伝的に分化した2つの遺伝系統(系統Iと系統II)が検出された.系統I(赤色)はサハリンから本州の中部山岳まで広域に分布するのに対し,系統II(青色)は本州だけに分布する.これらの遺伝系統の分布域は本州で広く重複するが,系統Iは北海道と東北に多く,系統IIは本州中西部に多い傾向がみられた(図2).
系統Iと系統IIが分岐した年代は,氷期と間氷期が繰り返された第四紀の更新世(約258万年前から1万年前)であると推定された(図2).続いて,それぞれの系統の祖先が分布していたと考えられる地域を推定した.すると,系統Iと系統IIの共通祖先の分布域は,サハリンから本州中部山岳までの比較的まで広範囲であると推定され,特定の地域には限定されなかった.一方では,系統Iの祖先が分布していた地域はサハリンから本州北部(東北地方の北部)に,系統IIは本州中部(東北地方の南部から中部地方)の可能性が高いと推定された(図2).またmtDNA COI領域に基づき,一定のエリア内における遺伝的な分化度(すなわち遺伝的多様度)の高い地域を可視化したところ,系統Iでは本州よりもサハリンや北海道で高く,系統IIでは本州北部よりも本州中部で遺伝的多様度が高い地域として評価された.
これらの結果は,更新世に繰り返されてきた氷期と間氷期サイクルの中で分布域の拡大・縮小を繰り返すうちに集団が分断化し,異なる地域で遺伝的な変異を蓄積したことを意味する.では,それぞれの系統の進化史の差異はどのようなものか? また,どのような要因で差異がもたらされたのか? このような疑問の下,系統間の遺伝構造の差異と,生息環境(標高)の差異の関係性について検討した.
サハリントビケラにおける分子系統解析
ミトコンドリアDNA(mtDNA)COI領域(648-bp),16S rRNA領域(417-bp)を結合した塩基配列に基づく系統解析を実施した結果,サハリントビケラの種内には別種として扱われてもよいレベルに大きく遺伝的に分化した2つの遺伝系統(系統Iと系統II)が検出された.系統I(赤色)はサハリンから本州の中部山岳まで広域に分布するのに対し,系統II(青色)は本州だけに分布する.これらの遺伝系統の分布域は本州で広く重複するが,系統Iは北海道と東北に多く,系統IIは本州中西部に多い傾向がみられた(図2).
系統Iと系統IIが分岐した年代は,氷期と間氷期が繰り返された第四紀の更新世(約258万年前から1万年前)であると推定された(図2).続いて,それぞれの系統の祖先が分布していたと考えられる地域を推定した.すると,系統Iと系統IIの共通祖先の分布域は,サハリンから本州中部山岳までの比較的まで広範囲であると推定され,特定の地域には限定されなかった.一方では,系統Iの祖先が分布していた地域はサハリンから本州北部(東北地方の北部)に,系統IIは本州中部(東北地方の南部から中部地方)の可能性が高いと推定された(図2).またmtDNA COI領域に基づき,一定のエリア内における遺伝的な分化度(すなわち遺伝的多様度)の高い地域を可視化したところ,系統Iでは本州よりもサハリンや北海道で高く,系統IIでは本州北部よりも本州中部で遺伝的多様度が高い地域として評価された.
これらの結果は,更新世に繰り返されてきた氷期と間氷期サイクルの中で分布域の拡大・縮小を繰り返すうちに集団が分断化し,異なる地域で遺伝的な変異を蓄積したことを意味する.では,それぞれの系統の進化史の差異はどのようなものか? また,どのような要因で差異がもたらされたのか? このような疑問の下,系統間の遺伝構造の差異と,生息環境(標高)の差異の関係性について検討した.
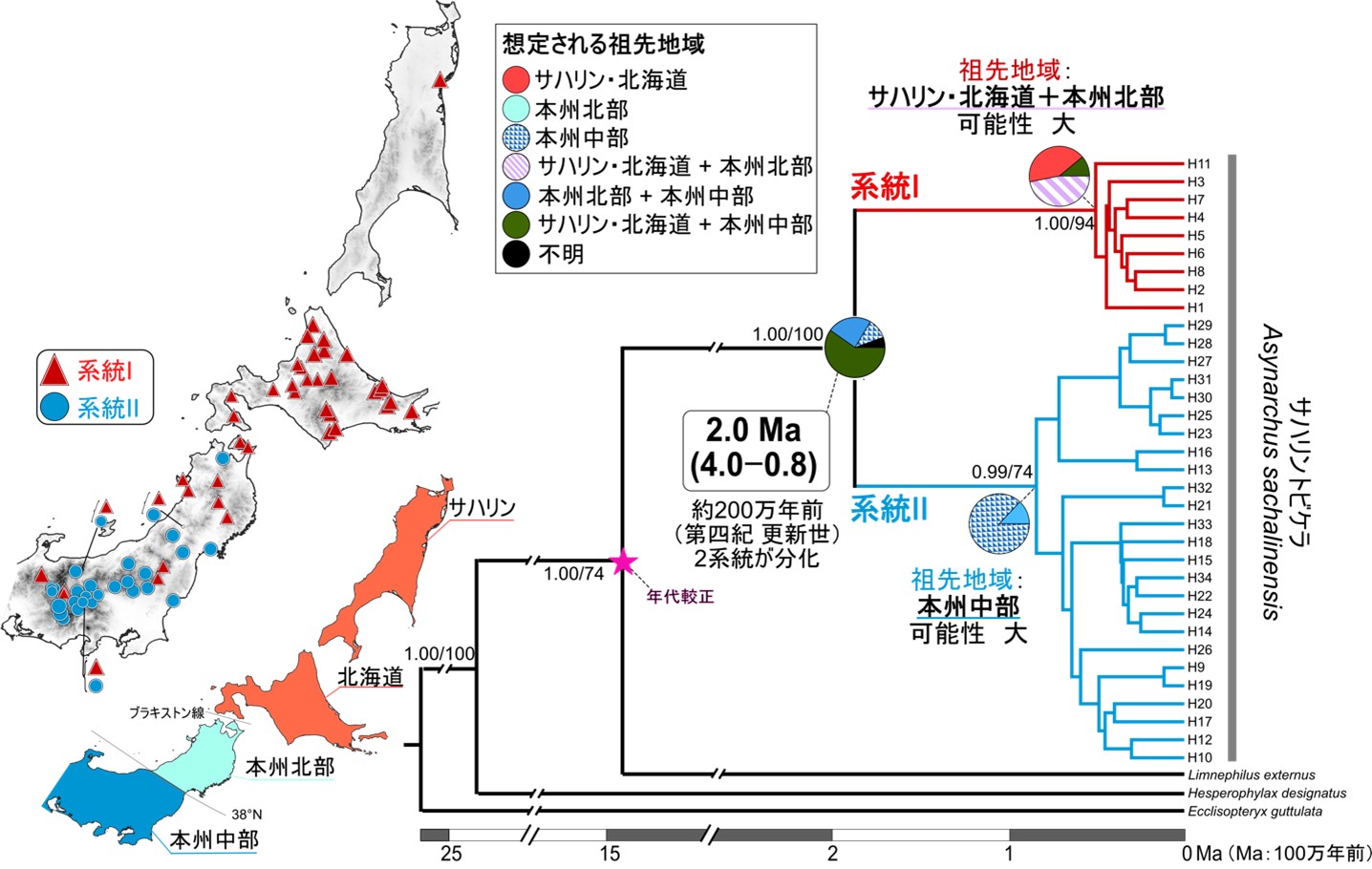
図2.サハリントビケラの採集地点と系統解析結果.ミトコンドリアDNA(mtDNA)COI領域(648 bp)と16S rRNA(417 bp)領域に基づく系統解析の結果(ベイズ系統樹),サハリントビケラは遺伝的に大きく分化した2つの系統から構成されることが判明した(系統I・系統II)また,これらの分岐年代を推定した結果,これらの系統の分化は約200万年前(第四紀 更新世)と評価され,氷期–間氷期サイクルの影響が示唆された.祖先地域推定の結果,系統Iの祖先はサハリン・北海道+本州北部に棲息した可能性が高く推定され,系統IIの祖先は本州中部に生息した可能性が高く推定された.なお生息地は3つの地域(多くの生物種で種や系統の分布境界となるブラキストン線,北緯38度線を境界)に分け,その組み合わせで祖先地域の推定が実施された.
サハリントビケラの遺伝構造の違いと生息地の標高の違い
系統Iと系統IIのそれぞれについて,mtDNA COI領域のハプロタイプネットワーク*4を作成したところ,系統Iは優占的なハプロタイプ(C2)を中心とする「花火型」のネットワーク構造を示した(図3).このような「花火型」のネットワーク構造は,近年の急激な集団サイズの拡大が生じたことを示す.一方で,系統IIは系統Iとは異なり,優占的なハプロタイプは検出されず,花火型のネットワークとはならなかった(図3).このようなネットワーク構造は,長期にわたり安定して集団サイズを維持したことを示す.また,検出されたハプロタイプの種類と割合を地図上に示したところ,系統Iは優占的なハプロタイプであるC2が分布域広域から検出された(図3).一方,系統IIは広域で検出されるハプロタイプは存在せず,地域で固有のハプロタイプが複数存在した.特に,ハプロタイプC26は乗鞍岳の高山帯だけから検出され,他集団がもつハプロタイプから4塩基の差異を有していた.(図3).両系統を構成する個体が採取された標高を調べたところ,系統Iに対して系統IIが,より高い標高帯から採取されていることが明らかとなった(図3).
系統Iと系統IIのそれぞれについて,mtDNA COI領域のハプロタイプネットワーク*4を作成したところ,系統Iは優占的なハプロタイプ(C2)を中心とする「花火型」のネットワーク構造を示した(図3).このような「花火型」のネットワーク構造は,近年の急激な集団サイズの拡大が生じたことを示す.一方で,系統IIは系統Iとは異なり,優占的なハプロタイプは検出されず,花火型のネットワークとはならなかった(図3).このようなネットワーク構造は,長期にわたり安定して集団サイズを維持したことを示す.また,検出されたハプロタイプの種類と割合を地図上に示したところ,系統Iは優占的なハプロタイプであるC2が分布域広域から検出された(図3).一方,系統IIは広域で検出されるハプロタイプは存在せず,地域で固有のハプロタイプが複数存在した.特に,ハプロタイプC26は乗鞍岳の高山帯だけから検出され,他集団がもつハプロタイプから4塩基の差異を有していた.(図3).両系統を構成する個体が採取された標高を調べたところ,系統Iに対して系統IIが,より高い標高帯から採取されていることが明らかとなった(図3).
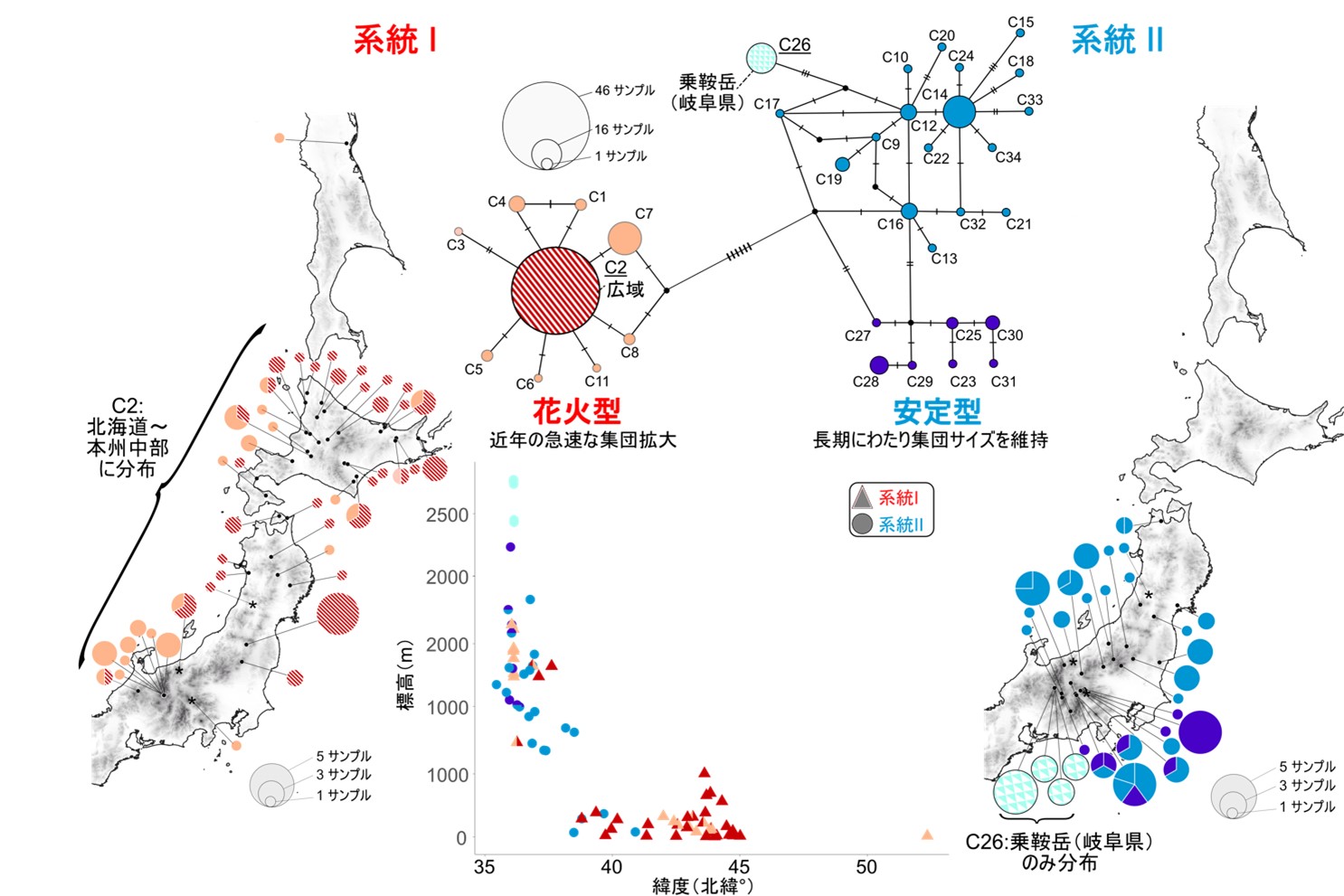
図3.サハリントビケラのハプロタイプネットワークと地理的分布,生息地の緯度・標高.ミトコンドリアDNA(mtDNA)COI領域(648 bp)に基づくハプロタイプネットワーク,ハプロタイプの地理的分布.系統Iのハプロタイプネットワーク(左)はハプロタイプC2を中心とした花火型を示し,C2は分布域広域から検出された.系統IIのハプロタイプネットワークは花火型を形成せず,地域固有のハプロタイプが多く検出された(例:C26は乗鞍岳の高山帯だけから採取).採集地点の緯度と標高の関係から(中央),系統Iは北方・低地に多くみられ,系統IIは南方・高地に多くみられた.つまり,系統IとIIには,遺伝構造,生息地の標高に違いがみられた.(なお,図中の各シンボルの配色はハプロタイプの配色と対応している)
サハリントビケラが好むのは冷涼な池沼や泉である.系統Iが多く検出された本州北部や北海道では,このような環境が比較的低地に連続的に存在することが多い.その場合,移動・分散は起こりやすくなり,他の集団との遺伝的交流も容易となる.そのため,地域に固有のハプロタイプは検出されにくい.一方,系統IIが中心的に分布する本州・中部山岳では,冷涼な池沼や泉が山岳の高地に孤立・散在的に存在する.つまり,山岳の起伏に富んだ地形が分散障壁となり,他の集団との遺伝的交流の機会が制限される.そのため地域固有のハプロタイプが検出されたと推察される.
これまで,高山帯では,ヨツバシオガマやミネズオウなどといった高山植物を中心に,生物系統地理・集団遺伝学的解析が進められ,中部山岳域にはそれぞれの生物種が長期にわたり集団を維持できる環境が存在した可能性が示されてきた.今回我々が対象とした水生昆虫は,幼虫期を水中で過ごす.水中は極端な低温や乾燥を防ぐことができるため,氷期にも凍結しなかった山岳内の泉(湧水)で生き延び,遺伝的変異を蓄積しつつ,長期にわたって集団が維持されてきた可能性がある.従って,これまでに知られてきた日本列島の山岳に生息する陸上生物とは異なる進化史のパターンが示唆された.つまり,サハリントビケラは更新世の氷期–間氷期の最中,分布域を拡大・縮小するうちに集団が分化し,一方は移動・分散が容易な北方の低地の池沼や泉に,もう一方は南方の高地の孤立・散在的な池や泉に生息するようになったと推測される.
今回の研究では,「孤立的な高地」と,「連続的な低地」という異なる生息環境に適応する系統間で遺伝構造を比較したことで,生息環境が遺伝的分化に影響することが強く示された.そして,環境の違いは生き物の生態的な違いをもたらす可能性が高い.また,山岳には気温差や地形の起伏など,様々な移動・分散障壁(遺伝子流動の制限)が存在する.地理的に近接する集団間のごく最近の遺伝子流動が検出できる鋭敏な遺伝子マーカー(ゲノムワイド*5な一塩基多型(SNPs)*6やマイクロサテライト*7など)を用いた遺伝子解析により,地理的に隔離された山塊内の集団間の遺伝子流動の有無を顕著に検出できる.
特に今回の研究では,高山帯の集団が同一山塊(乗鞍岳)の集団とは遺伝的に大きく離れていることが示された.この遺伝的差異をもたらす要因は,高山帯特有の環境や地形が寄与している可能性が高い.今後,生態研究や鋭敏な遺伝子マーカーを用いた研究を行い,高山帯における種分化要因の追究をきっかけに,山岳が果たす生物多様性創出のメカニズムの追究を行う予定である.
また,高山帯(系統II)と亜高山帯(系統I)の集団間での大きな遺伝的分化がみられた乗鞍岳については,高山帯・亜高山帯それぞれ4箇所ずつの池沼を対象に,2022年に,年間を通した生活史調査と高山帯−亜高山帯間での遺伝子流動を評価する研究を実施した.これらの成果については,今回の論文とは独立した形で別論文として投稿しているが,高山帯−亜高山帯の集団間で生活史が大きく異なっていることや,ゲノムワイドなSNPs解析においても長年にわたる遺伝子流動が生じていないことが明らかとなっている.すなわち,生殖的隔離も成立していることを示しており,種分化の条件(新種としての条件)を満たしていることが明らかとなった.
これまで,高山帯では,ヨツバシオガマやミネズオウなどといった高山植物を中心に,生物系統地理・集団遺伝学的解析が進められ,中部山岳域にはそれぞれの生物種が長期にわたり集団を維持できる環境が存在した可能性が示されてきた.今回我々が対象とした水生昆虫は,幼虫期を水中で過ごす.水中は極端な低温や乾燥を防ぐことができるため,氷期にも凍結しなかった山岳内の泉(湧水)で生き延び,遺伝的変異を蓄積しつつ,長期にわたって集団が維持されてきた可能性がある.従って,これまでに知られてきた日本列島の山岳に生息する陸上生物とは異なる進化史のパターンが示唆された.つまり,サハリントビケラは更新世の氷期–間氷期の最中,分布域を拡大・縮小するうちに集団が分化し,一方は移動・分散が容易な北方の低地の池沼や泉に,もう一方は南方の高地の孤立・散在的な池や泉に生息するようになったと推測される.
今回の研究では,「孤立的な高地」と,「連続的な低地」という異なる生息環境に適応する系統間で遺伝構造を比較したことで,生息環境が遺伝的分化に影響することが強く示された.そして,環境の違いは生き物の生態的な違いをもたらす可能性が高い.また,山岳には気温差や地形の起伏など,様々な移動・分散障壁(遺伝子流動の制限)が存在する.地理的に近接する集団間のごく最近の遺伝子流動が検出できる鋭敏な遺伝子マーカー(ゲノムワイド*5な一塩基多型(SNPs)*6やマイクロサテライト*7など)を用いた遺伝子解析により,地理的に隔離された山塊内の集団間の遺伝子流動の有無を顕著に検出できる.
特に今回の研究では,高山帯の集団が同一山塊(乗鞍岳)の集団とは遺伝的に大きく離れていることが示された.この遺伝的差異をもたらす要因は,高山帯特有の環境や地形が寄与している可能性が高い.今後,生態研究や鋭敏な遺伝子マーカーを用いた研究を行い,高山帯における種分化要因の追究をきっかけに,山岳が果たす生物多様性創出のメカニズムの追究を行う予定である.
また,高山帯(系統II)と亜高山帯(系統I)の集団間での大きな遺伝的分化がみられた乗鞍岳については,高山帯・亜高山帯それぞれ4箇所ずつの池沼を対象に,2022年に,年間を通した生活史調査と高山帯−亜高山帯間での遺伝子流動を評価する研究を実施した.これらの成果については,今回の論文とは独立した形で別論文として投稿しているが,高山帯−亜高山帯の集団間で生活史が大きく異なっていることや,ゲノムワイドなSNPs解析においても長年にわたる遺伝子流動が生じていないことが明らかとなっている.すなわち,生殖的隔離も成立していることを示しており,種分化の条件(新種としての条件)を満たしていることが明らかとなった.
【用語解説】
*1 ハビタット: 生物の生息場所のこと.ただし,その生物が生息するために必要な,温度や湿度などの非生物的な要因や,餌や競争相手など他の生物との関わり合い(生物的な要因)も含めた概念.
*2 リフュージア: 広い範囲で生物が絶滅するような環境下(今回は氷期)で,生物が生き残ることができた場所.「逃避地」ともいう.
*3 トビケラ: チョウ類やガ類に近い昆虫で,幼虫は芋虫型で,ほとんどの種が水中で生活する.口から吐く糸で筒状の巣を形成し,巣内で生活する種も多い.成虫は陸上生活するが,翅には(チョウ類やガ類がもつような)鱗粉はなく,その代わりに細かい毛が生えている.
*4 ハプロタイプネットワーク: 同一の遺伝子座位におけるDNAの塩基配列の違いに基づき区別された遺伝子型のうち,ミトコンドリアの遺伝子型をハプロタイプといい,配列の類似性に基づきハプロタイプ同士を線で繋いだ「ネットワーク」として表現したものをいう.図3における各円は検出された各ハプロタイプを表し,ハプロタイプ同士をつなぐ線にクロスする線の本数はハプロタイプ間の塩基置換数(ハプロタイプの差異)である.この本数が多いほど,ハプロタイプ間の差異は大きくなる.
*5 ゲノムワイド: 生物の性質を決定する遺伝情報をゲノムといい,ゲノムはDNAの塩基配列ならなる.その生物がもつ全ゲノムを網羅するような解析を実施することを「ゲノムワイドな解析」として表現される.
*6 一塩基多型(SNPs): 生物のもつDNAの塩基配列が一塩基単位で変異した違い.この変異はゲノム中に多く存在し,とても近縁な遺伝的関係(親と子など)の間にも違いがあることが知られている.一塩基多型(SNPs)の変異を調べることで,ミトコンドリアDNAの塩基配列では区別することができなかった,近接する集団同士の遺伝的な違いの検出も可能となっている.近年では,ゲノムワイドなSNPs解析などが多用されている.
*7 マイクロサテライト: 生物のDNAの塩基配列の中に存在する,数個の塩基が複数回繰り返した配列のこと.一塩基多型(SNPs)同様,非常に近い遺伝的関係の間で違いがあるため,近接する集団間の遺伝的な違いを検出できる.
【備考】
高山帯に関する調査は環境省許可番号 環中中国許2109012, 2206271のもと実施.
【論文タイトルと著者等】
タイトル:Evolutionary history of a cold-adapted limnephilid caddisfly: Effects of climate change and Topography on Genetic Structure
著者:Hirohisa SUZUKI, Masaki TAKENAKA, Koji TOJO
掲載誌:Molecular Phylogenetics and Evolution 191: 107967
掲載日:2024年2月公開(オンラインでは1月19日に公開)
URL:https://doi.org/10.1016/j.ympev.2023.107967
D O I:10.1016/j.ympev.2023.107967
著者:
Hirohisa SUZUKI 鈴木啓久(信州大学大学院 博士課程 社会人大学院生)筆頭著者
Masaki TAKENAKA 竹中將起(信州大学)
Koji TOJO 東城幸治(信州大学)責任著者
*1 ハビタット: 生物の生息場所のこと.ただし,その生物が生息するために必要な,温度や湿度などの非生物的な要因や,餌や競争相手など他の生物との関わり合い(生物的な要因)も含めた概念.
*2 リフュージア: 広い範囲で生物が絶滅するような環境下(今回は氷期)で,生物が生き残ることができた場所.「逃避地」ともいう.
*3 トビケラ: チョウ類やガ類に近い昆虫で,幼虫は芋虫型で,ほとんどの種が水中で生活する.口から吐く糸で筒状の巣を形成し,巣内で生活する種も多い.成虫は陸上生活するが,翅には(チョウ類やガ類がもつような)鱗粉はなく,その代わりに細かい毛が生えている.
*4 ハプロタイプネットワーク: 同一の遺伝子座位におけるDNAの塩基配列の違いに基づき区別された遺伝子型のうち,ミトコンドリアの遺伝子型をハプロタイプといい,配列の類似性に基づきハプロタイプ同士を線で繋いだ「ネットワーク」として表現したものをいう.図3における各円は検出された各ハプロタイプを表し,ハプロタイプ同士をつなぐ線にクロスする線の本数はハプロタイプ間の塩基置換数(ハプロタイプの差異)である.この本数が多いほど,ハプロタイプ間の差異は大きくなる.
*5 ゲノムワイド: 生物の性質を決定する遺伝情報をゲノムといい,ゲノムはDNAの塩基配列ならなる.その生物がもつ全ゲノムを網羅するような解析を実施することを「ゲノムワイドな解析」として表現される.
*6 一塩基多型(SNPs): 生物のもつDNAの塩基配列が一塩基単位で変異した違い.この変異はゲノム中に多く存在し,とても近縁な遺伝的関係(親と子など)の間にも違いがあることが知られている.一塩基多型(SNPs)の変異を調べることで,ミトコンドリアDNAの塩基配列では区別することができなかった,近接する集団同士の遺伝的な違いの検出も可能となっている.近年では,ゲノムワイドなSNPs解析などが多用されている.
*7 マイクロサテライト: 生物のDNAの塩基配列の中に存在する,数個の塩基が複数回繰り返した配列のこと.一塩基多型(SNPs)同様,非常に近い遺伝的関係の間で違いがあるため,近接する集団間の遺伝的な違いを検出できる.
【備考】
高山帯に関する調査は環境省許可番号 環中中国許2109012, 2206271のもと実施.
【論文タイトルと著者等】
タイトル:Evolutionary history of a cold-adapted limnephilid caddisfly: Effects of climate change and Topography on Genetic Structure
著者:Hirohisa SUZUKI, Masaki TAKENAKA, Koji TOJO
掲載誌:Molecular Phylogenetics and Evolution 191: 107967
掲載日:2024年2月公開(オンラインでは1月19日に公開)
URL:https://doi.org/10.1016/j.ympev.2023.107967
D O I:10.1016/j.ympev.2023.107967
著者:
Hirohisa SUZUKI 鈴木啓久(信州大学大学院 博士課程 社会人大学院生)筆頭著者
Masaki TAKENAKA 竹中將起(信州大学)
Koji TOJO 東城幸治(信州大学)責任著者